障害による神経回路の再編と機能の回復
2017/06/19
上野 将紀
(新潟大学脳研究所 システム脳病態学分野)
email:上野将紀
領域融合レビュー, 6, e003 (2017) DOI: 10.7875/leading.author.6.e003
Masaki Ueno: Rewiring of neural circuits and functional recovery following brain and spinal cord injuries.

外傷や血管障害により脳や脊髄が障害をうけると,神経回路は破綻し重篤な機能不全を起こす.成体は神経回路を新たに再建する能力にとぼしいため,脳や脊髄の障害に対し根本的な治療法はないのが現状である.しかし,近年の研究により,残存した神経回路が限定的ではありながら再編を起こし,障害ののちに機能が変容することがわかってきた.このレビューにおいては,これまでに明らかにされてきた神経回路の再編の機序について,げっ歯類における研究を中心に解説する.神経回路の再編の過程を理解しその質と量とを高めることにより,その機能の回復をもたらす戦略がみえてくるものと考えられる.
脳や脊髄は感覚,認知,運動といった機能によりわれわれの行動を規定し,また,自律神経により体内の環境を維持する,まさに中枢となる器官である.脳や脊髄には無数の神経回路が精巧にはりめぐらされておりその機能をはたす.血管障害や外傷といった要因により脳や脊髄が障害をうける可能性は身近なものであり,わが国では脳血管障害による患者が約120万人,脳挫傷や脊髄損傷の患者も約7万人いるとされる.これらの要因によりいちど神経回路が壊れると機能不全や麻痺が起こり社会生活が困難になるため,神経回路の機能を向上させる治療法の開発がもとめられている.機能の回復には失われた神経回路を再建することが必要であるが,成体の脳は神経回路を新たに構築する能力は非常にとぼしい.これまで,神経の再生能を上昇させようとする試みは多くなされてきたが,脳における神経回路の複雑さゆえ,再建に有効な手立てはいまだ確立されていない.
一方で,失われた神経回路の機能は障害ののち一定の程度の自然回復を示したり,あるいは逆に,増悪したりと変容することが古くから知られていた.近年の膨大な研究成果から,こうした機能の変化は障害からのがれて残存した神経回路の可塑的かつ代償的な変化,すなわち“再編”により起こることがわかってきた.成体の脳においても神経回路が変化しうることから,残された神経回路を生かした治療法が開発される希望がみえてきた.このレビューにおいては,障害により起こる神経回路の再編に関する基礎研究の取り組みおよびその成果について解説し,そこから,機能の回復をみちびくための展望および課題について模索する.
脳に可塑性のポテンシャルのあることは古くから知られていたが,それが脳や脊髄が障害されたのちの機能の回復の神経基盤としてきちんと観察されるようになったのは近年になってからであろう.サルにおける一連の実験は,そののちの神経回路の再編の研究に弾みをつけるきっかけのひとつとなった1)(図1a).大脳皮質の1次運動野には体のそれぞれの部位を動かす運動領域がある.これらは,大脳皮質の局所に電気刺激をあたえておのおのの部位の運動反応をみることによりわかる.ここでたとえば,手指や前腕に対応する領域に脳梗塞を起こすと,支配領域の消失にともない運動機能が障害される.脳梗塞ののちに手指に運動訓練を施すと,梗塞巣の周囲の残った領域において手指や前腕の動きをになう領域が拡大し,それにともない機能も回復した1).残った神経回路が可塑的に変化して機能の回復に寄与しうることが示唆されたのである.では,この過程において,脳ではいったい何が起こっているのであろうか.
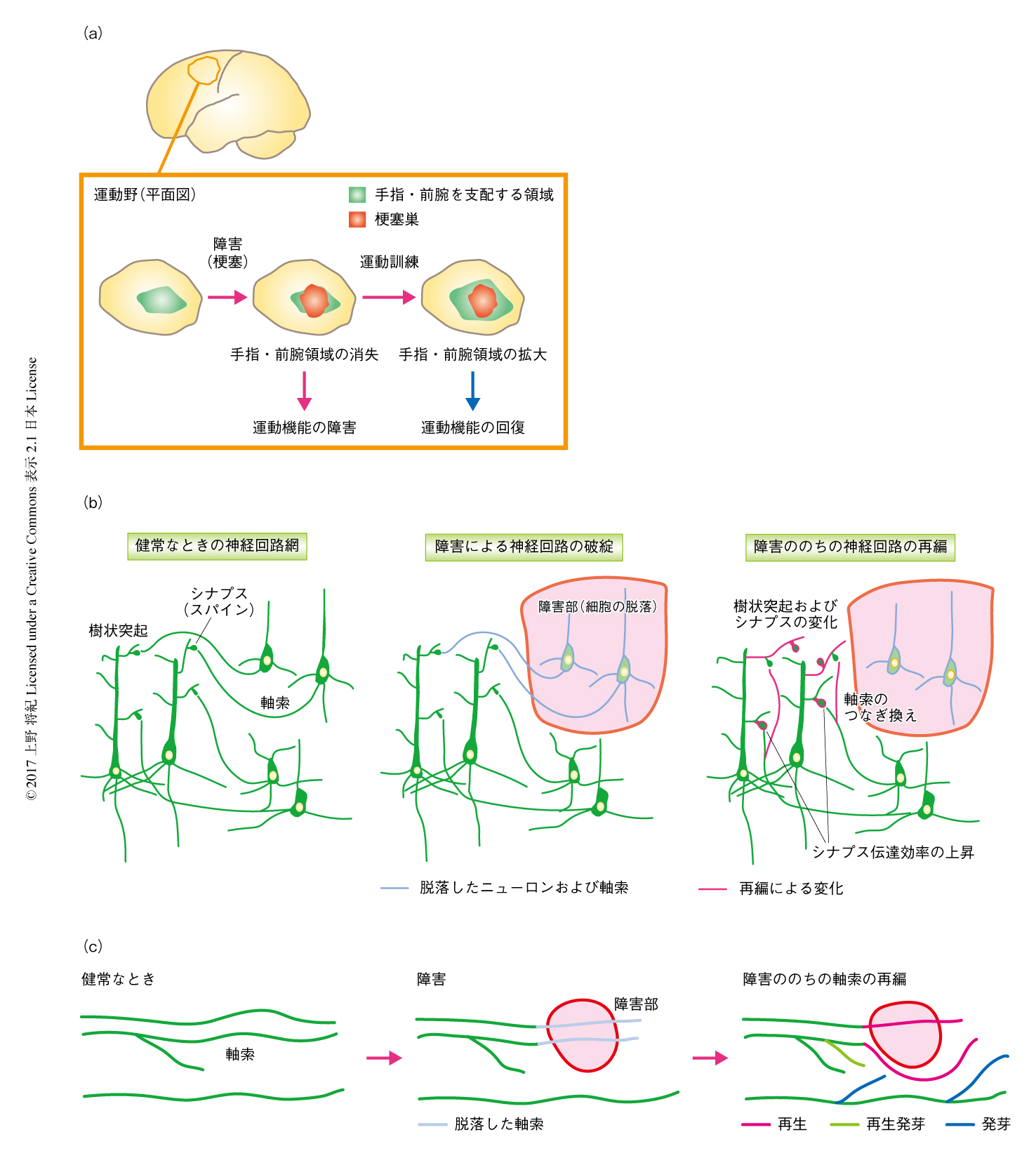
変化のもとになったのは神経回路の接続の再編と考えられる.これまでの研究により,ニューロンの軸索や樹状突起,シナプスの接続,さらには,神経伝達効率の変化など,さまざまなレベルの可塑的な変化が神経回路の再編にかかわることがわかってきた2)(図1b).たとえば,2光子顕微鏡を用いて梗塞の近傍のニューロンを観察すると,樹状突起はダイナミックに変化し,梗塞から数週間のあいだ,シナプス入力をうけるスパインのターンオーバーが増大するという3).こうした細胞体およびスパインの再編には接続する相手が必要なことから,軸索の側にも変化が起こるとされ,これらの変化が神経回路のつなぎ換えおよび機能の変化をもたらすと考えられる.実際に,軸索を介した再編は多くの研究において観察され,形態の変化をともなうもっともダイナミックな変化であるといえる.このレビューにおいては,とくに軸索が示す挙動を中心として神経回路の再編の過程について述べる.
軸索の再編には,大きく分けて以下の3つの種類がある(図1c).いちど切断された軸索から軸索が伸長する“再生”(regeneration),切断された軸索の途中から新たな軸索の側枝が生じる“再生発芽”(regenerative sprouting),障害をうけていない軸索から新たな側枝が伸長する“発芽”(sprouting),である4,5).中枢神経系において軸索の再生が自然に起こることはほぼない.すでに1世紀以上もまえ,スペインの神経解剖学者Cajalは,神経の再生について実験と思考を重ね,中枢神経系において再生の泉は枯渇していると結論づけた6).そこでは,切断された末梢神経は再生するが脊髄には侵入できないことを観察し,中枢神経には軸索の伸長に適さない環境のあることを予見した.そののち,近年にいたるまで,さまざまなくふうにより人為的に再生がうながされてきたが,神経回路を再建するには長い距離を伸長する必要もあり,いまもって,再生は機能の回復をもたらすには難易度が高いと考えられている.一方で,発芽については,実験的にある特定の軸索や領域を障害すると周囲の残った軸索から発芽が起こることが報告されている.また,そうして形成された神経回路は機能を変化させる力のあることもわかってきている.よって,発芽は神経回路の再編において重要な要素のひとつであると考えられつつある.
では実際に,発芽はどのような状況で起こるのだろうか.外傷や脳血管障害においては運動をつかさどる神経回路および機能がしばしば障害され生活を困難にすることが問題になっている.これらの理由から,神経再編の研究領域においては,運動回路の再編や再生の機序がよく探求されてきた.運動回路は大脳皮質や脳幹部といった上位の中枢と,筋への出力系になる下位の脊髄からなる.中枢からの指令は軸索の下行路により脊髄へと伝達される.なかでも,大脳皮質と脊髄とを結び自発運動や巧緻運動に重要とされる皮質脊髄路は,ヒトにおいてもその障害が運動不全のおもな要因になることから広く研究の対象になっている(図2a).
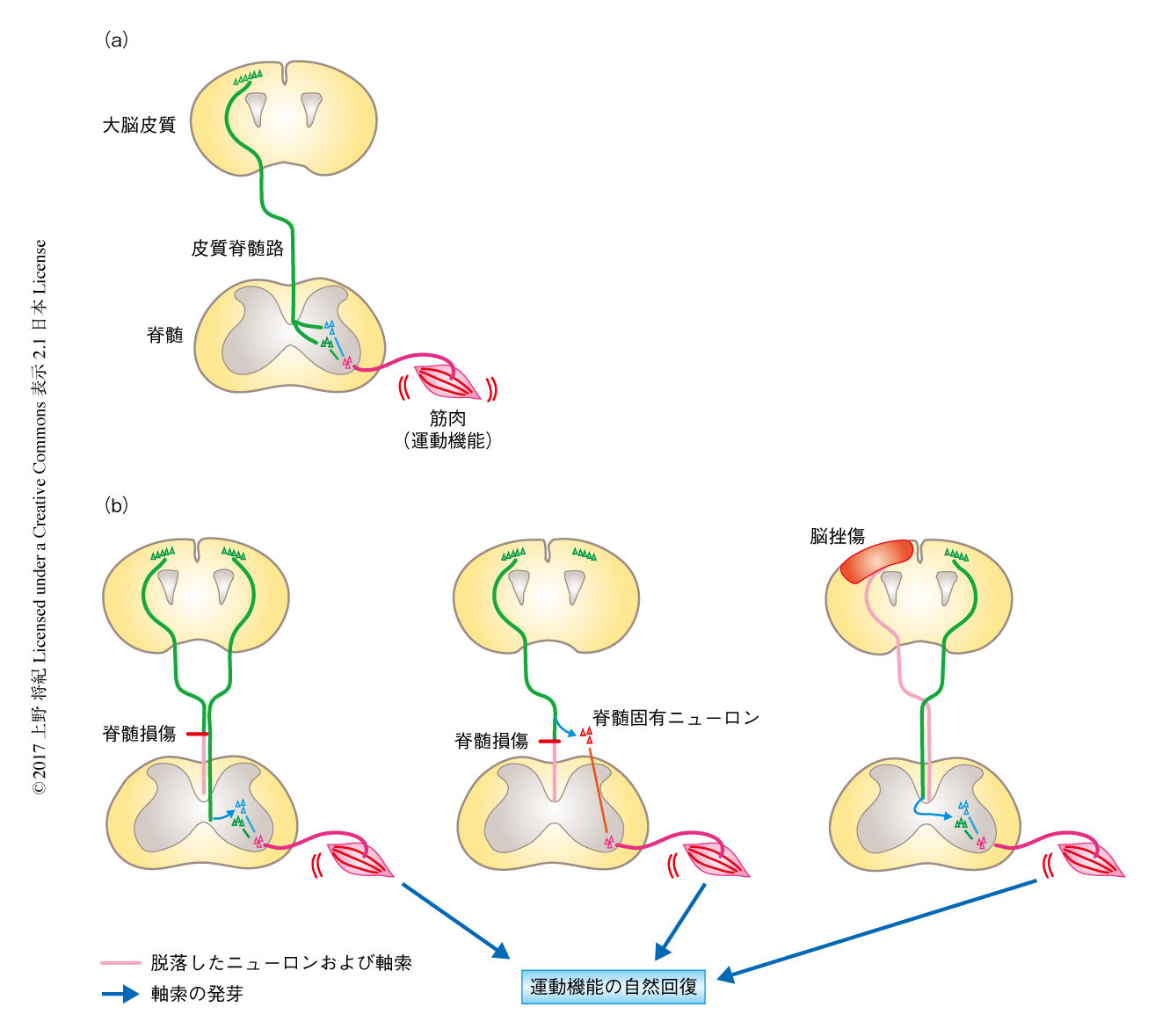
この皮質脊髄路を構成するニューロンや軸索が障害をうけると,残った神経回路が発芽により再編することが多くの研究において示されている.多くの場合,発芽により形成された神経回路が機能の自然な回復に貢献するようである.たとえば,ラットにおいて,脊髄の損傷ののち残存した皮質脊髄路が発芽により代償性の神経回路を形成し,それを実験的に切断すると回復した機能がふたたび低下したことから,新たに形成された神経回路が運動機能の回復に寄与することが実証された7)(図2b,左).また,同じくラットにおいて,脊髄の損傷により皮質脊髄路が切断されると軸索の途中から側枝が伸長し,損傷部の上位と下位とを結びつける脊髄固有ニューロンに接続しなおし,この接続は損傷部をこえて後肢の機能を支配する腰髄へのバイパスを形成し後肢の機能の回復をもたらすことが示された8)(図2b,中).
筆者らの研究グループは,脳挫傷モデルにおいて皮質脊髄路の可塑的な変化を観察した9)(図2b,右).マウスにおいて片側の大脳皮質の運動野を広範に損傷すると皮質脊髄路が脱落する.皮質脊髄路の軸索は脳幹部において交差して脊髄へと伸長するため,損傷と反対側の前肢の機能が障害された.しかし,数週間たつと運動機能は徐々に自然回復した.損傷部と反対側の残存した皮質脊髄路を調べたところ,頸髄において多数の発芽が起こっていた.これらの軸索は正中をこえて反対側へと伸長して脊髄のニューロンと接続し,新たに大脳皮質から筋肉へと連なる神経回路を形成していた.実際に,非損傷側の大脳皮質の運動野を電気刺激すると麻痺側の前肢の筋運動が起こり,また,この神経回路を切断すると自然回復した運動機能がふたたび失われた.この結果から,残った皮質脊髄路が新たな神経回路を形成し運動機能の自然回復に寄与することがわかった.
こうした神経回路の再編は,げっ歯類のみならず,ヒトに近いサルにおいても顕著に起こることが示されている.たとえば,頸髄半切断損傷モデルにおいては,損傷部の下位の残った皮質脊髄路において発芽により軸索の密度が上昇し,シナプス結合部位の増加など構造的な変化が起こるという10).また,こうした解剖学的な変化は上肢の機能の回復の程度とよく相関していた.げっ歯類およびサルと発芽が種に広く保存された現象であることから,実際にヒトの患者においても発芽は起こりうるのではないだろうか.
発芽による再編は,脳や脊髄における障害の場所や大きさにより,皮質脊髄路にとどまらず,皮質網様体路や網様体脊髄路11),皮質赤核路12) といったほかの上下行路にも起こることが報告されている.また,障害の近傍の領域にも起こる.たとえば,脳梗塞においては大脳皮質の運動野や感覚野など運動機能に重要な脳領野のあいだの接続も障害される.大脳皮質の局所に梗塞を起こすと大脳皮質のネットワークにおいても発芽が起こり,梗塞巣の周囲に残存したニューロンにおいて新たな接続が増加する13).似たような結合様式の変化は,サルの大脳梗塞モデルにおいても認められている14).こうした軸索の変化による再編が,機能マップの再編1) の一端となることが推測される.
また,神経回路の再編は運動回路にとどまらず,感覚回路15-17) や自律神経回路18) にも起こる.たとえば,交感神経回路には脳幹部など上位の中枢から胸髄へとむかう下行路があり,そこから末梢神経によりおのおのの臓器へとシグナルが伝達される.脊髄に損傷を起こすと下行路への入力の遮断にともない自律神経に障害が起こることが知られている.筆者らは,損傷部の下位に残存した交感神経回路が興奮性ニューロンとの接続を増加させて変化し,逆に神経回路を活性化することを見い出した18).神経回路の再編は,障害ののち,脳や脊髄において普遍的に起こる現象なのである.
再編された神経回路は機能の回復に貢献できることがみえてきたが,この再編をうながす機構は何であろうか.この機序がわかれば,神経回路の形成の過程をうまく制御して有効な治療法へと結びつきそうである.
神経回路の一部が障害されると周囲の残った軸索が新たに侵入するという現象は,じつは古くから知られている.1940年代には,末梢において皮膚へと伸びている末梢神経を切断するとほかの領域から軸索が侵入することが示されている19).こうした軸索の変化は,そののち,軸索を標識する技術の進歩により,中枢神経系においても徐々に鮮明になった.1958年には,標的とする軸索を切断し変性した軸索を染色するNauta法の登場により,ネコにおいて,脊髄へと伸長する下行路や感覚神経を障害し長い期間がたつと残った感覚神経の線維が新たに脊髄へ侵入することが観察された20).1969年には,ラットの中隔核において,海馬あるいは視床下部から伸びる中隔核への入力のいずれかを遮断すると,もう一方の軸索が中隔核のニューロンと新たにシナプスにより神経回路を形成することが電子顕微鏡により示された21).同様に1973年には,海馬において嗅内皮質からの入力がなくなると,交連線維が侵入して新たな神経回路が形成されることが電気生理学的に示された22).軸索を中心とした再編により新たに神経回路が形成されることが明らかにされたわけである.当時はまだ,形成された神経回路が生理学的な機能にまで昇華するかどうかはよくわかっていなかったが,成体の脳において障害ののちに神経回路が変化しうることの重要性はすでに認識されていた22,23).
この再編を起こす機序については,じつはいまもってその全貌は明らかにされていない.おそらく,神経回路における障害が引き金となり,それに反応して周囲の軸索が伸長し標的となるニューロンと接続するのであろう.こうした神経回路の形成は,そもそも,発生や発達の段階において起こるものであるから,そこでの現象がヒントとなると考えられてきた.神経回路の発達の過程においては,軸索が神経栄養因子や軸索ガイダンス因子に導かれて正しく投射し,さらに,経験にもとづく神経活動に依存的なシグナルにより神経回路が精緻化する.そのような過程は障害をうけた脳にも起こるのだろうか.
軸索が発芽や枝分かれを起こす詳細な機構についてはまだよくわかっていないが,発達期に軸索の誘引,成長,枝分かれを起こす因子はいくつか知られている.障害をうけたときには,たとえば,軸索からの入力を失ったニューロンになんらかの変化が生じ,そこから放出されるこうした因子が別の軸索を誘引する可能性が考えられる(図3,左上).たとえば,再編をうながすタンパク質の探索のため遺伝子発現スクリーニングが試みられている24,25).これらの研究においては,下行路を障害したのちに脊髄において神経栄養因子や軸索ガイダンス因子の増加が認められているが,軸索の再編をうながす責任タンパク質の同定にまではいたっていない.筆者らは,脳挫傷モデルにおいて,神経栄養因子や軸索ガイダンス因子を網羅的にスクリーニングした結果,標的となる脊髄のニューロンには神経栄養因子BDNFが発現しており,障害ののちの皮質脊髄路の軸索の枝分かれに必要であることを明らかにした9).これだけでは再編の過程のすべては説明しきれなかったが,再編の分子機構の探究に光をあてたといえる.そののち,軸索の入力を失った脊髄の領域において栄養因子CNTFの発現が上昇することが見い出され,それが軸索の発芽をひき起こすことが示唆された26).また,大脳皮質においては梗塞ののちに神経栄養因子や軸索ガイダンス因子の発現が上昇し,梗塞部の周囲のニューロンが発現するGDF10により軸索の発芽がうながされることが見い出されている13,27).また,古くから,軸索の入力を失った領域においてシナプスの近傍のアストロサイトにも形態的な変化の起こることが知られている23).その役割についてはよくわかっていなかったが,近年,アストロサイトから放出されるトロンボスポンジン1およびトロンボスポンジン2が梗塞ののちの軸索の発芽およびシナプスの新生にかかわることが報告されている28).

ここで述べたような因子が協同的に再編をうながすのか,あるいは,個々の障害の状況により使われる因子が変わるのかは,いまのところよくわかっていない.未知の因子の探索も含め,再編の普遍的な機構の解明が待たれるところである.
ここまで述べた栄養および誘引物質以外の可能性として,軸索どうしの競合作用をもとにした機序も考えられる23)(図3,右上).この競合作用は,もともと神経回路の発達の過程においてみられるものである.軸索が標的となる領域において均一に分布するため,別個のニューロンから伸長した軸索どうしが反発し整列する.障害をうけると軸索の脱落によりこの競合は解除され,その結果,空いた空間にほかの軸索が入り込むという現象が起こる可能性が考えられる.その真偽はまだ定かではないが,皮質脊髄路どうしなど同種の軸索の競合である場合もあれば9),皮質脊髄路と感覚神経など別種の軸索どうしの場合があるかもしれない20,29).ただし,軸索の脱落のない領域において再編が起こるパターンもあり,その場合,脱競合の機構はそぐわないかもしれない.
最後に,発達の過程においては,大まかな神経回路が形成されたのち,神経活動に依存的して接続が精緻化される.この過程においては,Hebb則により出力側と入力側の神経活動の一致した接続が残され,必要のない軸索やシナプスは刈り込まれる.障害ののちには軸索が選択的あるいはランダムに標的となる領域へと入り込むが,この再編の過程においても,最終的にはHebb則により類似の機能をもつ軸索と標的となるニューロンとの接続が残ると考えられる(図3,下).リハビリテーションにより特定の機能をもつ神経回路が優勢になるという効果をみると,神経活動にそった接続の強化が起こっていそうである.筆者らも,解剖学的に,障害により皮質脊髄路の発芽が亢進したのち,刈り込みにより軸索が減少することを見い出した9).こうした生理学的あるいは解剖学的な変化は,あたかも神経回路の発達の過程を再現するかのようである.
神経回路が再編できることはわかってきたものの,この変化は莫大に起こるものではなく,失われた神経回路を補いきるものではない.では,再編を大幅に増加させることは可能なのだろうか.神経回路が再建されないおもな要因は,まず障害により多くのニューロンが失われることにあるが,残存した神経回路を有効に使おうとした場合,Cajalが予見したように,中枢神経において軸索が容易に伸長できないことにも要因があろう.以来,ながらくつづいてきた研究の結果,成体の脳において軸索が伸長あるいは再生できない要因は外因性および内因性の2つに大別されてきた.
外因性の要因として,当初,再生に適さない中枢神経の環境においてはニューロンの成長を促進する因子が不足していると考えられた(図4).発達の過程において豊富に存在する神経栄養因子や軸索ガイダンス因子は,成体の脳においては異なる発現の様式を示したり発現量が減少していたりする.実際,障害ののちに神経栄養因子であるNT-3やBDNFをくわえると軸索の伸長が亢進する9,30).したがって,神経栄養因子の添加は再編をうながす有益な戦略のひとつかもしれない.ただ,こうした神経栄養因子がほかの神経回路も刺激してしまい異常な神経回路が形成される危険性もある.この場合には正常な機能をさまたげるため,適切な神経回路の再編を起こすには神経栄養因子に反応する神経回路の特異性が問題になるであろう.軸索ガイダンス因子も成体の脳においてはもはや発達期と同様の発現はみられない.しかし,成体においても再編された軸索はまだ発現している軸索ガイダンス因子にしたがいルートを選択できることから31),軸索の誘導の機序として使える可能性は残る.いずれにしても,発達の段階におけるこれらの因子の発現を再現して神経回路の再編を進めることはなかなか困難といえる.
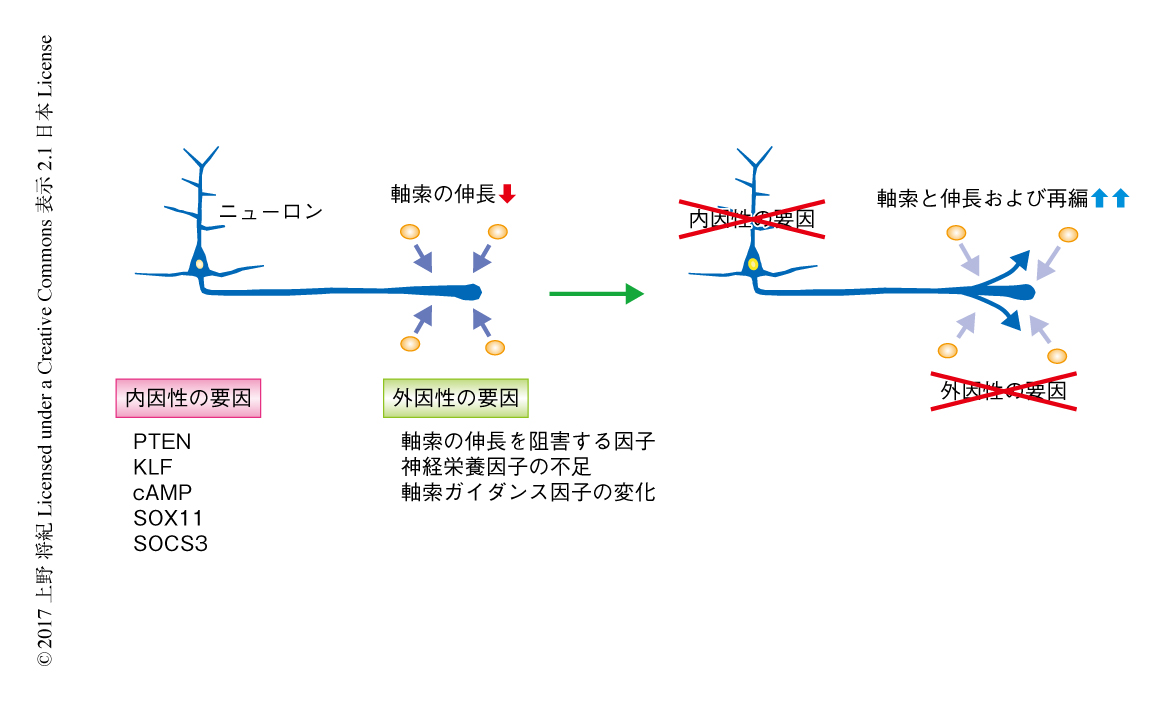
1980年代に入ると,逆に,成体の中枢神経には軸索の伸長を阻害する外因性の因子の存在することが示されはじめた32).これまで,このような外来性の因子としてミエリンに発現するタンパク質がみつかっている(図4).ミエリンの存在により軸索の伸長が阻害されることが明らかにされてから33),今日までに,その責任タンパク質であるNogoや多くの関連タンパク質また受容体が見い出されている5).これらのタンパク質は軸索の成長円錐の崩壊を介して軸索の伸長を抑制するとされる.実際に,脳や脊髄が障害されたときにこれらのタンパク質を阻害することにより,再生が促進されるかどうかはいまだ議論があるが,発芽は促進される5,34,35).こうした軸索の伸長を阻害する因子の探索やそのシグナルの研究は現在進行形でつづいている.阻害する方法やその量,それにともなうほかの神経回路への副作用など,考えるべき点は多いが,いまのところ,神経回路の再編を亢進するために非常に期待される標的であるといえる.
もうひとつの柱が内因性の要因である(図4).脳の発達期において障害により一部の神経回路が失われると,残存した神経回路が成体の脳に比べ大幅な変化を示すことは古くから知られている.筆者らも,マウスにおいて脳の障害により起こる皮質脊髄路の再編は生後14日までは大規模に起こることを示した36).こうした再編の規模の変化は,ひとつには脳の発達の途中から現われるミエリンをはじめとする軸索の伸長を阻害する因子の増加,すなわち,外因性の要因によるものとされているが,もうひとつは,ニューロンそれ自体のもつ軸索の伸長能が成体になるにしたがい減衰することによることがわかっている.これが内因性の要因である.実際に,網膜のニューロンのもつ軸索の伸長能は,生後,劇的に低下するという37).その原因の探究はいまも進められているところだが,たとえば,転写因子KLF4は細胞が成熟するにつれて発現が上昇し,それが軸索の伸長能の低下に関連している38).また逆に,同じファミリーの転写因子KLF7は若いニューロンに多く発現し成体においては減少するが,脊髄の損傷ののちに過剰に発現させると皮質脊髄路の再生および発芽が促進される39).そのほかにも,cAMP濃度40),Sox11 41),SOCS3 26) といった内在性の要因が軸索の伸長力を規定する.こうした要因のなかでおそらくもっとも効果をあげているのはPtenのノックアウトであろう42).ニューロンの成熟や障害が起こると細胞の成長をになうmTORの活性が低下するが,mTORを阻害するホスファターゼであるPtenをノックアウトしたところ,mTORとともに翻訳の亢進をはじめとする細胞におけるシグナルが活性化され,皮質脊髄路の再生および発芽が顕著に促進された.ニューロンあるいは軸索の上限のない成長は神経の機能に負にはたらく可能性もあり,シグナルを活性化する時期や量について慎重な検討が必要ではあるが,内因性のシグナルの活性化による戦略のすぐれたポテンシャルをみせた研究であるといえる.
最後に,ニューロンや神経回路それ自体の神経活動も,神経回路の形成をうながすための重要な要因である.発達の過程において,神経活動は軸索の枝分かれなどの伸長および神経回路の精緻化の両方にはたらく.これは,障害ののちにもみられ,たとえば,皮質脊髄路を継続的に電気刺激すると発芽による再編が増加し機能の回復にも貢献するという43,44).この神経活動がどのような機序により軸索の伸長を促進するのか,その詳細はわかっていないが,内因性のシグナルの活性化がかかわると考えられており,再編を亢進するよい手段と期待されている.気をつけるべきは,適切な神経回路の形成には,こうした外来からの人為的な刺激をするにせよ,行動に即した神経活動の惹起が必要であることである.同期した神経活動が発芽に必要との報告もあり45),リハビリテーションなど,より自然な神経活動の惹起との組合せを考慮せねばならない.
神経回路の再編を人為的に増加させる研究が成果をあげつつある一方,伸長した軸索をいかに標的となる適切なニューロンと接続させるかが課題として残っている.障害ののち,機能が代償的な行動へと変化したり,あるいは,必要な機能が逆に阻害や増悪したりするような不適応(maladaptive)な変化を示すことがある.これらの現象にも再編した神経回路が影響している可能性が高く,その場合には,機能の発揮に不適切な接続の形成が根幹にあると考えられる.可塑性はどのような方向へも機能を変容させる力をもつわけである.
たとえば,あるひとつのかたよった運動トレーニングにより神経回路の再編をうながすと,ひとつの運動試験による機能が回復を示すとしても,別の運動試験による機能が異常になることが報告されている46).筆者らも,リハビリテーションの違いによる神経回路の再編への影響について検討したが,マウスにおいて,エサをとる単純なリーチ動作をくり返し訓練するとその動作しか回復の効果が得られなかった47).逆に,両手足を協調的に動かす幅広い訓練をすると広範な運動機能への効果が認められ,また,皮質脊髄路の再編への効果も高かった.“不使用の学習”(learned non-use)といわれる現象もこれに関連すると思われ,使用しないと神経回路は機能しない方向へと収束してしまう.サルの基礎実験から確立されたCI(constraint induced)療法はこの点興味深い48),健常な側を拘束し麻痺の側の手を強制的に使用させると運動機能の回復が得られるという.実際,げっ歯類において同様のCI療法を施すと,機能の回復にともない皮質脊髄路や皮質赤核路の再編が亢進すると報告されている12,25).これらは,リハビリテーションの効果の神経基盤の一端を示すものであるが,同時に,もとめられる機能にそった適切な神経回路を形成するよう訓練しなければならないことを暗示している.
適切な刺激で介入することにより神経回路をうまく形成できればいいが,もともと神経回路に内在する機構がはたらいてしまうと,機能の回復にそうようにうまく再編が進まない場合も多い.たとえば,皮質脊髄路が障害により脱落すると,脱落した脊髄の領域に末梢からの固有感覚線維が侵入してしまい,その結果,筋反射が亢進してしまう29).この予期しない再編が筋の緊張が亢進する痙縮(spasticity)の要因のひとつになるとみられる.ほかにも,再編した神経回路が異常な接続を形成する場合,痛みや急激な血圧の上昇をひき起こす自律神経過反射にいたることも指摘されている18,49).また,人為的に過剰な再編を起こそうとする場合にも注意が必要である.たとえば,Ptenのノックアウトにより軸索の伸長が大きく増加しても機能の回復は顕著には起こらない50).Sox11の過剰な発現により皮質脊髄路の再生および発芽は増加するが,巧緻運動の機能はむしろ悪化する41).脊髄を損傷したのち神経栄養因子BDNFを損傷部およびその近傍に発現させると網様体脊髄路の軸索の伸長が増加するが,逆に,運動機能は低下し痙縮が増加してしまう51).
こうした事例をみると,機能の回復をもたらすには適切な量および適切な接続による再編が必要であることがわかる.近年,この点において画期的な成果が報告されている52).脳梗塞ののちしばらく抗Nogo-A抗体の投与により軸索の伸長を増加させる時期を設定し,のちに,リハビリテーションを施すという治療プロトコールを検討した結果,脊髄における皮質脊髄路の再編が亢進し,それとともに機能の回復の効果が高まった.逆に,抗Nogo-A抗体の投与とリハビリテーションを同時に行うと機能への効果は打ち消された.この場合にも皮質脊髄路の再編は亢進したが,軸索が異所性に過剰に侵入する像が観察され異常な神経回路が形成されたと推察された.脳の発達の過程においては,最初に軸索の伸長が起こり,そののち,神経活動により神経回路が精緻化する.この神経回路の発達を模した再編をひき起こす戦略として,この研究は興味深い結果を提示している.適切な時期および量をもって介入し正しい神経回路の接続をうながさねばならないのである.
神経回路の再編の研究における大きなゴールは,いかに神経回路の再編を制御し効果的に機能を再建できるか,その手法を見い出すことにある.そのためには,神経回路の再編の機序は何か,どのような方法が再編を増加させるか,再編した神経回路はどのように機能するか,といった基本的な課題を地道に解明することが必要である.とくに,再編された神経回路と発揮された機能とがどのように結びつくのかを実証することが,技術的な面も含め,これからの大きな課題のひとつといえる.これまでの研究により,樹状突起,スパイン,軸索の形態的な変化や生理学的な反応の変化が明らかにされ,神経回路が再編する実体がうかびあがっているが,そうした神経回路における現象がどのようにして機能と結びつくのか,結合の様式や動作原理の面から理解することが必要となっている.
脳の機能の理解は,そもそも,健常な脳においてもまだまだ解明の余地が大きい.正常な神経回路がどのような接続をもって機能するのか,そうした神経回路がどのような機序により形成されるのかについてもわかっていないことが多い.しかしながら,近年の神経科学分野の技術の進歩はめざましい.健常な脳のコネクトーム解析およびそれらの神経回路が織りなす機能の解析はいま隆盛をきわめているが,それらの知見をもとにすれば,障害をうけた脳の神経回路は健常な状態からどのように変化して機能しているのかより鮮明になるだろう.こうした技術は正常な脳やその発達における解析に用いられてきたが,障害をうけた脳の神経回路の解析にも徐々に使われはじめている.遺伝子改変技術53-55),光遺伝学および化学遺伝学的な手法18,26,52,55-57),ウイルストレーサー12,18,26,52,53),イメージング技術3,17) により,特定のニューロン種や神経回路を標的にした観察や機能の修飾が可能になっている.それにより,どのような神経回路が機能の回復あるいは増悪に結びつくのかが少しずつみえはじめている.また,分子機序からみると,再編された神経回路の形成の機構を知ることが今後の課題のひとつであろう.RNA-seq法をはじめとした大規模な遺伝子解析の技術がそれに貢献するかもしれない.
げっ歯類モデルは,こうした最先端の技術を使って特異的な神経回路にアクセスしやすいことから,これからも広く用いられるであろう.一方で,こうした基礎研究の成果が臨床応用へとつながる例はほんのひとにぎりであり,ヒトとのシステムの違いが効果の違いを生み出す可能性を十分に考えねばならない.種差を念頭に入れながら,神経回路のもつ根本的な機構を探求することが必要だろう.このレビューにおいては,げっ歯類を中心とした基礎研究について紹介したが,さきに述べた技術はサルにおける応用も試みられており,さまざまな制約が取り除かれればヒトに近い実験モデルによる研究も一段と進歩するであろう.脳の神経回路が自力で回復するしくみを知りそれを補助する方法を確立することが,脳神経疾患の治療法の開発やリハビリテーションの科学的な基盤の構築に貢献するものと期待される.
略歴:2006年 東京大学大学院農学生命科学研究科 修了,理化学研究所脳科学総合研究センター,千葉大学大学院医学研究院,大阪大学大学院医学系研究科,米国Cincinnati Children's Hospital Medical Center,科学技術振興機構さきがけ研究者を経て,2016年より新潟大学脳研究所 特任教授.
研究テーマ:健常な脳および障害をうけた脳における神経回路の構成とその動作原理.
© 2017 上野 将紀 Licensed under CC 表示 2.1 日本
(新潟大学脳研究所 システム脳病態学分野)
email:上野将紀
領域融合レビュー, 6, e003 (2017) DOI: 10.7875/leading.author.6.e003
Masaki Ueno: Rewiring of neural circuits and functional recovery following brain and spinal cord injuries.
要 約
外傷や血管障害により脳や脊髄が障害をうけると,神経回路は破綻し重篤な機能不全を起こす.成体は神経回路を新たに再建する能力にとぼしいため,脳や脊髄の障害に対し根本的な治療法はないのが現状である.しかし,近年の研究により,残存した神経回路が限定的ではありながら再編を起こし,障害ののちに機能が変容することがわかってきた.このレビューにおいては,これまでに明らかにされてきた神経回路の再編の機序について,げっ歯類における研究を中心に解説する.神経回路の再編の過程を理解しその質と量とを高めることにより,その機能の回復をもたらす戦略がみえてくるものと考えられる.
はじめに
脳や脊髄は感覚,認知,運動といった機能によりわれわれの行動を規定し,また,自律神経により体内の環境を維持する,まさに中枢となる器官である.脳や脊髄には無数の神経回路が精巧にはりめぐらされておりその機能をはたす.血管障害や外傷といった要因により脳や脊髄が障害をうける可能性は身近なものであり,わが国では脳血管障害による患者が約120万人,脳挫傷や脊髄損傷の患者も約7万人いるとされる.これらの要因によりいちど神経回路が壊れると機能不全や麻痺が起こり社会生活が困難になるため,神経回路の機能を向上させる治療法の開発がもとめられている.機能の回復には失われた神経回路を再建することが必要であるが,成体の脳は神経回路を新たに構築する能力は非常にとぼしい.これまで,神経の再生能を上昇させようとする試みは多くなされてきたが,脳における神経回路の複雑さゆえ,再建に有効な手立てはいまだ確立されていない.
一方で,失われた神経回路の機能は障害ののち一定の程度の自然回復を示したり,あるいは逆に,増悪したりと変容することが古くから知られていた.近年の膨大な研究成果から,こうした機能の変化は障害からのがれて残存した神経回路の可塑的かつ代償的な変化,すなわち“再編”により起こることがわかってきた.成体の脳においても神経回路が変化しうることから,残された神経回路を生かした治療法が開発される希望がみえてきた.このレビューにおいては,障害により起こる神経回路の再編に関する基礎研究の取り組みおよびその成果について解説し,そこから,機能の回復をみちびくための展望および課題について模索する.
1.神経回路の再編と機能の回復
脳に可塑性のポテンシャルのあることは古くから知られていたが,それが脳や脊髄が障害されたのちの機能の回復の神経基盤としてきちんと観察されるようになったのは近年になってからであろう.サルにおける一連の実験は,そののちの神経回路の再編の研究に弾みをつけるきっかけのひとつとなった1)(図1a).大脳皮質の1次運動野には体のそれぞれの部位を動かす運動領域がある.これらは,大脳皮質の局所に電気刺激をあたえておのおのの部位の運動反応をみることによりわかる.ここでたとえば,手指や前腕に対応する領域に脳梗塞を起こすと,支配領域の消失にともない運動機能が障害される.脳梗塞ののちに手指に運動訓練を施すと,梗塞巣の周囲の残った領域において手指や前腕の動きをになう領域が拡大し,それにともない機能も回復した1).残った神経回路が可塑的に変化して機能の回復に寄与しうることが示唆されたのである.では,この過程において,脳ではいったい何が起こっているのであろうか.
変化のもとになったのは神経回路の接続の再編と考えられる.これまでの研究により,ニューロンの軸索や樹状突起,シナプスの接続,さらには,神経伝達効率の変化など,さまざまなレベルの可塑的な変化が神経回路の再編にかかわることがわかってきた2)(図1b).たとえば,2光子顕微鏡を用いて梗塞の近傍のニューロンを観察すると,樹状突起はダイナミックに変化し,梗塞から数週間のあいだ,シナプス入力をうけるスパインのターンオーバーが増大するという3).こうした細胞体およびスパインの再編には接続する相手が必要なことから,軸索の側にも変化が起こるとされ,これらの変化が神経回路のつなぎ換えおよび機能の変化をもたらすと考えられる.実際に,軸索を介した再編は多くの研究において観察され,形態の変化をともなうもっともダイナミックな変化であるといえる.このレビューにおいては,とくに軸索が示す挙動を中心として神経回路の再編の過程について述べる.
軸索の再編には,大きく分けて以下の3つの種類がある(図1c).いちど切断された軸索から軸索が伸長する“再生”(regeneration),切断された軸索の途中から新たな軸索の側枝が生じる“再生発芽”(regenerative sprouting),障害をうけていない軸索から新たな側枝が伸長する“発芽”(sprouting),である4,5).中枢神経系において軸索の再生が自然に起こることはほぼない.すでに1世紀以上もまえ,スペインの神経解剖学者Cajalは,神経の再生について実験と思考を重ね,中枢神経系において再生の泉は枯渇していると結論づけた6).そこでは,切断された末梢神経は再生するが脊髄には侵入できないことを観察し,中枢神経には軸索の伸長に適さない環境のあることを予見した.そののち,近年にいたるまで,さまざまなくふうにより人為的に再生がうながされてきたが,神経回路を再建するには長い距離を伸長する必要もあり,いまもって,再生は機能の回復をもたらすには難易度が高いと考えられている.一方で,発芽については,実験的にある特定の軸索や領域を障害すると周囲の残った軸索から発芽が起こることが報告されている.また,そうして形成された神経回路は機能を変化させる力のあることもわかってきている.よって,発芽は神経回路の再編において重要な要素のひとつであると考えられつつある.
では実際に,発芽はどのような状況で起こるのだろうか.外傷や脳血管障害においては運動をつかさどる神経回路および機能がしばしば障害され生活を困難にすることが問題になっている.これらの理由から,神経再編の研究領域においては,運動回路の再編や再生の機序がよく探求されてきた.運動回路は大脳皮質や脳幹部といった上位の中枢と,筋への出力系になる下位の脊髄からなる.中枢からの指令は軸索の下行路により脊髄へと伝達される.なかでも,大脳皮質と脊髄とを結び自発運動や巧緻運動に重要とされる皮質脊髄路は,ヒトにおいてもその障害が運動不全のおもな要因になることから広く研究の対象になっている(図2a).
この皮質脊髄路を構成するニューロンや軸索が障害をうけると,残った神経回路が発芽により再編することが多くの研究において示されている.多くの場合,発芽により形成された神経回路が機能の自然な回復に貢献するようである.たとえば,ラットにおいて,脊髄の損傷ののち残存した皮質脊髄路が発芽により代償性の神経回路を形成し,それを実験的に切断すると回復した機能がふたたび低下したことから,新たに形成された神経回路が運動機能の回復に寄与することが実証された7)(図2b,左).また,同じくラットにおいて,脊髄の損傷により皮質脊髄路が切断されると軸索の途中から側枝が伸長し,損傷部の上位と下位とを結びつける脊髄固有ニューロンに接続しなおし,この接続は損傷部をこえて後肢の機能を支配する腰髄へのバイパスを形成し後肢の機能の回復をもたらすことが示された8)(図2b,中).
筆者らの研究グループは,脳挫傷モデルにおいて皮質脊髄路の可塑的な変化を観察した9)(図2b,右).マウスにおいて片側の大脳皮質の運動野を広範に損傷すると皮質脊髄路が脱落する.皮質脊髄路の軸索は脳幹部において交差して脊髄へと伸長するため,損傷と反対側の前肢の機能が障害された.しかし,数週間たつと運動機能は徐々に自然回復した.損傷部と反対側の残存した皮質脊髄路を調べたところ,頸髄において多数の発芽が起こっていた.これらの軸索は正中をこえて反対側へと伸長して脊髄のニューロンと接続し,新たに大脳皮質から筋肉へと連なる神経回路を形成していた.実際に,非損傷側の大脳皮質の運動野を電気刺激すると麻痺側の前肢の筋運動が起こり,また,この神経回路を切断すると自然回復した運動機能がふたたび失われた.この結果から,残った皮質脊髄路が新たな神経回路を形成し運動機能の自然回復に寄与することがわかった.
こうした神経回路の再編は,げっ歯類のみならず,ヒトに近いサルにおいても顕著に起こることが示されている.たとえば,頸髄半切断損傷モデルにおいては,損傷部の下位の残った皮質脊髄路において発芽により軸索の密度が上昇し,シナプス結合部位の増加など構造的な変化が起こるという10).また,こうした解剖学的な変化は上肢の機能の回復の程度とよく相関していた.げっ歯類およびサルと発芽が種に広く保存された現象であることから,実際にヒトの患者においても発芽は起こりうるのではないだろうか.
発芽による再編は,脳や脊髄における障害の場所や大きさにより,皮質脊髄路にとどまらず,皮質網様体路や網様体脊髄路11),皮質赤核路12) といったほかの上下行路にも起こることが報告されている.また,障害の近傍の領域にも起こる.たとえば,脳梗塞においては大脳皮質の運動野や感覚野など運動機能に重要な脳領野のあいだの接続も障害される.大脳皮質の局所に梗塞を起こすと大脳皮質のネットワークにおいても発芽が起こり,梗塞巣の周囲に残存したニューロンにおいて新たな接続が増加する13).似たような結合様式の変化は,サルの大脳梗塞モデルにおいても認められている14).こうした軸索の変化による再編が,機能マップの再編1) の一端となることが推測される.
また,神経回路の再編は運動回路にとどまらず,感覚回路15-17) や自律神経回路18) にも起こる.たとえば,交感神経回路には脳幹部など上位の中枢から胸髄へとむかう下行路があり,そこから末梢神経によりおのおのの臓器へとシグナルが伝達される.脊髄に損傷を起こすと下行路への入力の遮断にともない自律神経に障害が起こることが知られている.筆者らは,損傷部の下位に残存した交感神経回路が興奮性ニューロンとの接続を増加させて変化し,逆に神経回路を活性化することを見い出した18).神経回路の再編は,障害ののち,脳や脊髄において普遍的に起こる現象なのである.
2.神経回路の再編の機序
再編された神経回路は機能の回復に貢献できることがみえてきたが,この再編をうながす機構は何であろうか.この機序がわかれば,神経回路の形成の過程をうまく制御して有効な治療法へと結びつきそうである.
神経回路の一部が障害されると周囲の残った軸索が新たに侵入するという現象は,じつは古くから知られている.1940年代には,末梢において皮膚へと伸びている末梢神経を切断するとほかの領域から軸索が侵入することが示されている19).こうした軸索の変化は,そののち,軸索を標識する技術の進歩により,中枢神経系においても徐々に鮮明になった.1958年には,標的とする軸索を切断し変性した軸索を染色するNauta法の登場により,ネコにおいて,脊髄へと伸長する下行路や感覚神経を障害し長い期間がたつと残った感覚神経の線維が新たに脊髄へ侵入することが観察された20).1969年には,ラットの中隔核において,海馬あるいは視床下部から伸びる中隔核への入力のいずれかを遮断すると,もう一方の軸索が中隔核のニューロンと新たにシナプスにより神経回路を形成することが電子顕微鏡により示された21).同様に1973年には,海馬において嗅内皮質からの入力がなくなると,交連線維が侵入して新たな神経回路が形成されることが電気生理学的に示された22).軸索を中心とした再編により新たに神経回路が形成されることが明らかにされたわけである.当時はまだ,形成された神経回路が生理学的な機能にまで昇華するかどうかはよくわかっていなかったが,成体の脳において障害ののちに神経回路が変化しうることの重要性はすでに認識されていた22,23).
この再編を起こす機序については,じつはいまもってその全貌は明らかにされていない.おそらく,神経回路における障害が引き金となり,それに反応して周囲の軸索が伸長し標的となるニューロンと接続するのであろう.こうした神経回路の形成は,そもそも,発生や発達の段階において起こるものであるから,そこでの現象がヒントとなると考えられてきた.神経回路の発達の過程においては,軸索が神経栄養因子や軸索ガイダンス因子に導かれて正しく投射し,さらに,経験にもとづく神経活動に依存的なシグナルにより神経回路が精緻化する.そのような過程は障害をうけた脳にも起こるのだろうか.
軸索が発芽や枝分かれを起こす詳細な機構についてはまだよくわかっていないが,発達期に軸索の誘引,成長,枝分かれを起こす因子はいくつか知られている.障害をうけたときには,たとえば,軸索からの入力を失ったニューロンになんらかの変化が生じ,そこから放出されるこうした因子が別の軸索を誘引する可能性が考えられる(図3,左上).たとえば,再編をうながすタンパク質の探索のため遺伝子発現スクリーニングが試みられている24,25).これらの研究においては,下行路を障害したのちに脊髄において神経栄養因子や軸索ガイダンス因子の増加が認められているが,軸索の再編をうながす責任タンパク質の同定にまではいたっていない.筆者らは,脳挫傷モデルにおいて,神経栄養因子や軸索ガイダンス因子を網羅的にスクリーニングした結果,標的となる脊髄のニューロンには神経栄養因子BDNFが発現しており,障害ののちの皮質脊髄路の軸索の枝分かれに必要であることを明らかにした9).これだけでは再編の過程のすべては説明しきれなかったが,再編の分子機構の探究に光をあてたといえる.そののち,軸索の入力を失った脊髄の領域において栄養因子CNTFの発現が上昇することが見い出され,それが軸索の発芽をひき起こすことが示唆された26).また,大脳皮質においては梗塞ののちに神経栄養因子や軸索ガイダンス因子の発現が上昇し,梗塞部の周囲のニューロンが発現するGDF10により軸索の発芽がうながされることが見い出されている13,27).また,古くから,軸索の入力を失った領域においてシナプスの近傍のアストロサイトにも形態的な変化の起こることが知られている23).その役割についてはよくわかっていなかったが,近年,アストロサイトから放出されるトロンボスポンジン1およびトロンボスポンジン2が梗塞ののちの軸索の発芽およびシナプスの新生にかかわることが報告されている28).
ここで述べたような因子が協同的に再編をうながすのか,あるいは,個々の障害の状況により使われる因子が変わるのかは,いまのところよくわかっていない.未知の因子の探索も含め,再編の普遍的な機構の解明が待たれるところである.
ここまで述べた栄養および誘引物質以外の可能性として,軸索どうしの競合作用をもとにした機序も考えられる23)(図3,右上).この競合作用は,もともと神経回路の発達の過程においてみられるものである.軸索が標的となる領域において均一に分布するため,別個のニューロンから伸長した軸索どうしが反発し整列する.障害をうけると軸索の脱落によりこの競合は解除され,その結果,空いた空間にほかの軸索が入り込むという現象が起こる可能性が考えられる.その真偽はまだ定かではないが,皮質脊髄路どうしなど同種の軸索の競合である場合もあれば9),皮質脊髄路と感覚神経など別種の軸索どうしの場合があるかもしれない20,29).ただし,軸索の脱落のない領域において再編が起こるパターンもあり,その場合,脱競合の機構はそぐわないかもしれない.
最後に,発達の過程においては,大まかな神経回路が形成されたのち,神経活動に依存的して接続が精緻化される.この過程においては,Hebb則により出力側と入力側の神経活動の一致した接続が残され,必要のない軸索やシナプスは刈り込まれる.障害ののちには軸索が選択的あるいはランダムに標的となる領域へと入り込むが,この再編の過程においても,最終的にはHebb則により類似の機能をもつ軸索と標的となるニューロンとの接続が残ると考えられる(図3,下).リハビリテーションにより特定の機能をもつ神経回路が優勢になるという効果をみると,神経活動にそった接続の強化が起こっていそうである.筆者らも,解剖学的に,障害により皮質脊髄路の発芽が亢進したのち,刈り込みにより軸索が減少することを見い出した9).こうした生理学的あるいは解剖学的な変化は,あたかも神経回路の発達の過程を再現するかのようである.
3.神経回路の再編をうながすには
神経回路が再編できることはわかってきたものの,この変化は莫大に起こるものではなく,失われた神経回路を補いきるものではない.では,再編を大幅に増加させることは可能なのだろうか.神経回路が再建されないおもな要因は,まず障害により多くのニューロンが失われることにあるが,残存した神経回路を有効に使おうとした場合,Cajalが予見したように,中枢神経において軸索が容易に伸長できないことにも要因があろう.以来,ながらくつづいてきた研究の結果,成体の脳において軸索が伸長あるいは再生できない要因は外因性および内因性の2つに大別されてきた.
外因性の要因として,当初,再生に適さない中枢神経の環境においてはニューロンの成長を促進する因子が不足していると考えられた(図4).発達の過程において豊富に存在する神経栄養因子や軸索ガイダンス因子は,成体の脳においては異なる発現の様式を示したり発現量が減少していたりする.実際,障害ののちに神経栄養因子であるNT-3やBDNFをくわえると軸索の伸長が亢進する9,30).したがって,神経栄養因子の添加は再編をうながす有益な戦略のひとつかもしれない.ただ,こうした神経栄養因子がほかの神経回路も刺激してしまい異常な神経回路が形成される危険性もある.この場合には正常な機能をさまたげるため,適切な神経回路の再編を起こすには神経栄養因子に反応する神経回路の特異性が問題になるであろう.軸索ガイダンス因子も成体の脳においてはもはや発達期と同様の発現はみられない.しかし,成体においても再編された軸索はまだ発現している軸索ガイダンス因子にしたがいルートを選択できることから31),軸索の誘導の機序として使える可能性は残る.いずれにしても,発達の段階におけるこれらの因子の発現を再現して神経回路の再編を進めることはなかなか困難といえる.
1980年代に入ると,逆に,成体の中枢神経には軸索の伸長を阻害する外因性の因子の存在することが示されはじめた32).これまで,このような外来性の因子としてミエリンに発現するタンパク質がみつかっている(図4).ミエリンの存在により軸索の伸長が阻害されることが明らかにされてから33),今日までに,その責任タンパク質であるNogoや多くの関連タンパク質また受容体が見い出されている5).これらのタンパク質は軸索の成長円錐の崩壊を介して軸索の伸長を抑制するとされる.実際に,脳や脊髄が障害されたときにこれらのタンパク質を阻害することにより,再生が促進されるかどうかはいまだ議論があるが,発芽は促進される5,34,35).こうした軸索の伸長を阻害する因子の探索やそのシグナルの研究は現在進行形でつづいている.阻害する方法やその量,それにともなうほかの神経回路への副作用など,考えるべき点は多いが,いまのところ,神経回路の再編を亢進するために非常に期待される標的であるといえる.
もうひとつの柱が内因性の要因である(図4).脳の発達期において障害により一部の神経回路が失われると,残存した神経回路が成体の脳に比べ大幅な変化を示すことは古くから知られている.筆者らも,マウスにおいて脳の障害により起こる皮質脊髄路の再編は生後14日までは大規模に起こることを示した36).こうした再編の規模の変化は,ひとつには脳の発達の途中から現われるミエリンをはじめとする軸索の伸長を阻害する因子の増加,すなわち,外因性の要因によるものとされているが,もうひとつは,ニューロンそれ自体のもつ軸索の伸長能が成体になるにしたがい減衰することによることがわかっている.これが内因性の要因である.実際に,網膜のニューロンのもつ軸索の伸長能は,生後,劇的に低下するという37).その原因の探究はいまも進められているところだが,たとえば,転写因子KLF4は細胞が成熟するにつれて発現が上昇し,それが軸索の伸長能の低下に関連している38).また逆に,同じファミリーの転写因子KLF7は若いニューロンに多く発現し成体においては減少するが,脊髄の損傷ののちに過剰に発現させると皮質脊髄路の再生および発芽が促進される39).そのほかにも,cAMP濃度40),Sox11 41),SOCS3 26) といった内在性の要因が軸索の伸長力を規定する.こうした要因のなかでおそらくもっとも効果をあげているのはPtenのノックアウトであろう42).ニューロンの成熟や障害が起こると細胞の成長をになうmTORの活性が低下するが,mTORを阻害するホスファターゼであるPtenをノックアウトしたところ,mTORとともに翻訳の亢進をはじめとする細胞におけるシグナルが活性化され,皮質脊髄路の再生および発芽が顕著に促進された.ニューロンあるいは軸索の上限のない成長は神経の機能に負にはたらく可能性もあり,シグナルを活性化する時期や量について慎重な検討が必要ではあるが,内因性のシグナルの活性化による戦略のすぐれたポテンシャルをみせた研究であるといえる.
最後に,ニューロンや神経回路それ自体の神経活動も,神経回路の形成をうながすための重要な要因である.発達の過程において,神経活動は軸索の枝分かれなどの伸長および神経回路の精緻化の両方にはたらく.これは,障害ののちにもみられ,たとえば,皮質脊髄路を継続的に電気刺激すると発芽による再編が増加し機能の回復にも貢献するという43,44).この神経活動がどのような機序により軸索の伸長を促進するのか,その詳細はわかっていないが,内因性のシグナルの活性化がかかわると考えられており,再編を亢進するよい手段と期待されている.気をつけるべきは,適切な神経回路の形成には,こうした外来からの人為的な刺激をするにせよ,行動に即した神経活動の惹起が必要であることである.同期した神経活動が発芽に必要との報告もあり45),リハビリテーションなど,より自然な神経活動の惹起との組合せを考慮せねばならない.
4.再編により適切な神経回路を形成するには
神経回路の再編を人為的に増加させる研究が成果をあげつつある一方,伸長した軸索をいかに標的となる適切なニューロンと接続させるかが課題として残っている.障害ののち,機能が代償的な行動へと変化したり,あるいは,必要な機能が逆に阻害や増悪したりするような不適応(maladaptive)な変化を示すことがある.これらの現象にも再編した神経回路が影響している可能性が高く,その場合には,機能の発揮に不適切な接続の形成が根幹にあると考えられる.可塑性はどのような方向へも機能を変容させる力をもつわけである.
たとえば,あるひとつのかたよった運動トレーニングにより神経回路の再編をうながすと,ひとつの運動試験による機能が回復を示すとしても,別の運動試験による機能が異常になることが報告されている46).筆者らも,リハビリテーションの違いによる神経回路の再編への影響について検討したが,マウスにおいて,エサをとる単純なリーチ動作をくり返し訓練するとその動作しか回復の効果が得られなかった47).逆に,両手足を協調的に動かす幅広い訓練をすると広範な運動機能への効果が認められ,また,皮質脊髄路の再編への効果も高かった.“不使用の学習”(learned non-use)といわれる現象もこれに関連すると思われ,使用しないと神経回路は機能しない方向へと収束してしまう.サルの基礎実験から確立されたCI(constraint induced)療法はこの点興味深い48),健常な側を拘束し麻痺の側の手を強制的に使用させると運動機能の回復が得られるという.実際,げっ歯類において同様のCI療法を施すと,機能の回復にともない皮質脊髄路や皮質赤核路の再編が亢進すると報告されている12,25).これらは,リハビリテーションの効果の神経基盤の一端を示すものであるが,同時に,もとめられる機能にそった適切な神経回路を形成するよう訓練しなければならないことを暗示している.
適切な刺激で介入することにより神経回路をうまく形成できればいいが,もともと神経回路に内在する機構がはたらいてしまうと,機能の回復にそうようにうまく再編が進まない場合も多い.たとえば,皮質脊髄路が障害により脱落すると,脱落した脊髄の領域に末梢からの固有感覚線維が侵入してしまい,その結果,筋反射が亢進してしまう29).この予期しない再編が筋の緊張が亢進する痙縮(spasticity)の要因のひとつになるとみられる.ほかにも,再編した神経回路が異常な接続を形成する場合,痛みや急激な血圧の上昇をひき起こす自律神経過反射にいたることも指摘されている18,49).また,人為的に過剰な再編を起こそうとする場合にも注意が必要である.たとえば,Ptenのノックアウトにより軸索の伸長が大きく増加しても機能の回復は顕著には起こらない50).Sox11の過剰な発現により皮質脊髄路の再生および発芽は増加するが,巧緻運動の機能はむしろ悪化する41).脊髄を損傷したのち神経栄養因子BDNFを損傷部およびその近傍に発現させると網様体脊髄路の軸索の伸長が増加するが,逆に,運動機能は低下し痙縮が増加してしまう51).
こうした事例をみると,機能の回復をもたらすには適切な量および適切な接続による再編が必要であることがわかる.近年,この点において画期的な成果が報告されている52).脳梗塞ののちしばらく抗Nogo-A抗体の投与により軸索の伸長を増加させる時期を設定し,のちに,リハビリテーションを施すという治療プロトコールを検討した結果,脊髄における皮質脊髄路の再編が亢進し,それとともに機能の回復の効果が高まった.逆に,抗Nogo-A抗体の投与とリハビリテーションを同時に行うと機能への効果は打ち消された.この場合にも皮質脊髄路の再編は亢進したが,軸索が異所性に過剰に侵入する像が観察され異常な神経回路が形成されたと推察された.脳の発達の過程においては,最初に軸索の伸長が起こり,そののち,神経活動により神経回路が精緻化する.この神経回路の発達を模した再編をひき起こす戦略として,この研究は興味深い結果を提示している.適切な時期および量をもって介入し正しい神経回路の接続をうながさねばならないのである.
おわりに
神経回路の再編の研究における大きなゴールは,いかに神経回路の再編を制御し効果的に機能を再建できるか,その手法を見い出すことにある.そのためには,神経回路の再編の機序は何か,どのような方法が再編を増加させるか,再編した神経回路はどのように機能するか,といった基本的な課題を地道に解明することが必要である.とくに,再編された神経回路と発揮された機能とがどのように結びつくのかを実証することが,技術的な面も含め,これからの大きな課題のひとつといえる.これまでの研究により,樹状突起,スパイン,軸索の形態的な変化や生理学的な反応の変化が明らかにされ,神経回路が再編する実体がうかびあがっているが,そうした神経回路における現象がどのようにして機能と結びつくのか,結合の様式や動作原理の面から理解することが必要となっている.
脳の機能の理解は,そもそも,健常な脳においてもまだまだ解明の余地が大きい.正常な神経回路がどのような接続をもって機能するのか,そうした神経回路がどのような機序により形成されるのかについてもわかっていないことが多い.しかしながら,近年の神経科学分野の技術の進歩はめざましい.健常な脳のコネクトーム解析およびそれらの神経回路が織りなす機能の解析はいま隆盛をきわめているが,それらの知見をもとにすれば,障害をうけた脳の神経回路は健常な状態からどのように変化して機能しているのかより鮮明になるだろう.こうした技術は正常な脳やその発達における解析に用いられてきたが,障害をうけた脳の神経回路の解析にも徐々に使われはじめている.遺伝子改変技術53-55),光遺伝学および化学遺伝学的な手法18,26,52,55-57),ウイルストレーサー12,18,26,52,53),イメージング技術3,17) により,特定のニューロン種や神経回路を標的にした観察や機能の修飾が可能になっている.それにより,どのような神経回路が機能の回復あるいは増悪に結びつくのかが少しずつみえはじめている.また,分子機序からみると,再編された神経回路の形成の機構を知ることが今後の課題のひとつであろう.RNA-seq法をはじめとした大規模な遺伝子解析の技術がそれに貢献するかもしれない.
げっ歯類モデルは,こうした最先端の技術を使って特異的な神経回路にアクセスしやすいことから,これからも広く用いられるであろう.一方で,こうした基礎研究の成果が臨床応用へとつながる例はほんのひとにぎりであり,ヒトとのシステムの違いが効果の違いを生み出す可能性を十分に考えねばならない.種差を念頭に入れながら,神経回路のもつ根本的な機構を探求することが必要だろう.このレビューにおいては,げっ歯類を中心とした基礎研究について紹介したが,さきに述べた技術はサルにおける応用も試みられており,さまざまな制約が取り除かれればヒトに近い実験モデルによる研究も一段と進歩するであろう.脳の神経回路が自力で回復するしくみを知りそれを補助する方法を確立することが,脳神経疾患の治療法の開発やリハビリテーションの科学的な基盤の構築に貢献するものと期待される.
文 献
- Nudo, R. J., Wise, B. M., SiFuentes, F. et al.: Neural substrates for the effects of rehabilitative training on motor recovery after ischemic infarct. Science, 272, 1791-1794 (1996)[PubMed]
- Murphy, T. H. & Corbett, D.: Plasticity during stroke recovery: from synapse to behaviour. Nat. Rev. Neurosci., 10, 861-872 (2009)[PubMed]
- Brown, C. E., Li, P., Boyd, J. D. et al.: Extensive turnover of dendritic spines and vascular remodeling in cortical tissues recovering from stroke. J. Neurosci., 27, 4101-4109 (2007)[PubMed]
- Tuszynski, M. H. & Steward, O.: Concepts and methods for the study of axonal regeneration in the CNS. Neuron, 74, 777-791 (2012)[PubMed]
- Geoffroy, C. G. & Zheng, B.: Myelin-associated inhibitors in axonal growth after CNS injury. Curr. Opin. Neurobiol., 27, 31-38 (2014)[PubMed]
- Ramon y Cajal, S.: Degeneration and Regeneration of the Nervous System. Oxford University Press, New York (1928)
- Weidner, N., Ner, A., Salimi, N. et al.: Spontaneous corticospinal axonal plasticity and functional recovery after adult central nervous system injury. Proc. Natl. Acad. Sci. USA, 98, 3513-3518 (2001)[PubMed]
- Bareyre, F. M., Kerschensteiner, M., Raineteau, O. et al.: The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat. Neurosci., 7, 269-277 (2004)[PubMed]
- Ueno, M., Hayano, Y., Nakagawa, H. et al.: Intraspinal rewiring of the corticospinal tract requires target-derived brain-derived neurotrophic factor and compensates lost function after brain injury. Brain, 135, 1253-1267 (2012)[PubMed]
- Rosenzweig, E. S., Courtine, G., Jindrich, D. L. et al.: Extensive spontaneous plasticity of corticospinal projections after primate spinal cord injury. Nat. Neurosci., 13, 1505-1510 (2010)[PubMed]
- Bachmann, L. C., Lindau, N. T., Felder, P. et al.: Sprouting of brainstem-spinal tracts in response to unilateral motor cortex stroke in mice. J. Neurosci., 34, 3378-3389 (2014)[PubMed]
- Ishida, A., Isa, K., Umeda, T. et al.: Causal link between the cortico-rubral pathway and functional recovery through forced impaired limb use in rats with stroke. J. Neurosci., 36, 455-467 (2016)[PubMed]
- Li, S., Overman, J. J., Katsman, D. et al.: An age-related sprouting transcriptome provides molecular control of axonal sprouting after stroke. Nat. Neurosci., 13, 1496-1504 (2010)[PubMed]
- Dancause, N., Barbay, S., Frost, S. B. et al.: Extensive cortical rewiring after brain injury. J. Neurosci., 25, 10167-10179 (2005)[PubMed]
- Hollis, E. R. 2nd, Ishiko, N., Pessian, M. et al.: Remodelling of spared proprioceptive circuit involving a small number of neurons supports functional recovery. Nat. Commun., 6, 6079 (2015)[PubMed]
- Brown, C. E., Aminoltejari, K., Erb, H. et al.: In vivo voltage-sensitive dye imaging in adult mice reveals that somatosensory maps lost to stroke are replaced over weeks by new structural and functional circuits with prolonged modes of activation within both the peri-infarct zone and distant sites. J. Neurosci., 29, 1719-1734 (2009)[PubMed]
- Takatsuru, Y., Fukumoto, D., Yoshitomo, M. et al.: Neuronal circuit remodeling in the contralateral cortical hemisphere during functional recovery from cerebral infarction. J. Neurosci., 29, 10081-10086 (2009)[PubMed]
- Ueno, M., Ueno-Nakamura, Y., Niehaus, J. et al.: Silencing spinal interneurons inhibits immune suppressive autonomic reflexes caused by spinal cord injury. Nat. Neurosci., 19, 784-787 (2016)[PubMed]
- Weddell, G., Guttmann, L. & Gutmann, E.: The local extension of nerve fibers into denervated areas of skin. J. Neurol. Psychiatry, 4, 206-225 (1941)[PubMed]
- Liu, C. N. & Chambers, W. W.: Intraspinal sprouting of dorsal root axons; development of new collaterals and preterminals following partial denervation of the spinal cord in the cat. AMA Arch. Neurol. Psychiatry, 79, 46-61 (1958)[PubMed]
- Raisman, G.: Neuronal plasticity in the septal nuclei of the adult rat. Brain Res., 14, 25-48 (1969)[PubMed]
- Lynch, G., Deadwyler, S. & Cotman, G.: Postlesion axonal growth produces permanent functional connections. Science, 180, 1364-1366 (1973)[PubMed]
- Raisman, G. & Field, P. M.: A quantitative investigation of the development of collateral reinnervation after partial deafferentation of the septal nuclei. Brain Res., 50, 241-264 (1973)[PubMed]
- Bareyre, F. M., Haudenschild, B. & Schwab, M. E.: Long-lasting sprouting and gene expression changes induced by the monoclonal antibody IN-1 in the adult spinal cord. J. Neurosci., 22, 7097-7110 (2002)[PubMed]
- Maier, I. C., Baumann, K., Thallmair, M. et al.: Constraint-induced movement therapy in the adult rat after unilateral corticospinal tract injury. J. Neurosci., 28, 9386-9403 (2008)[PubMed]
- Jin, D., Liu, Y., Sun, F. et al.: Restoration of skilled locomotion by sprouting corticospinal axons induced by co-deletion of PTEN and SOCS3. Nat. Commun., 6, 8074 (2015)[PubMed]
- Li, S., Nie, E. H., Yin, Y. et al.: GDF10 is a signal for axonal sprouting and functional recovery after stroke. Nat. Neurosci., 18, 1737-1745 (2015)[PubMed]
- Liauw, J., Hoang, S., Choi, M. et al.: Thrombospondins 1 and 2 are necessary for synaptic plasticity and functional recovery after stroke. J. Cereb. Blood Flow Metab., 28, 1722-1732 (2008)[PubMed]
- Tan, A. M., Chakrabarty, S., Kimura, H. et al.: Selective corticospinal tract injury in the rat induces primary afferent fiber sprouting in the spinal cord and hyperreflexia. J. Neurosci., 32, 12896-12908 (2012)[PubMed]
- Schnell, L., Schneider, R., Kolbeck, R. et al.: Neurotrophin-3 enhances sprouting of corticospinal tract during development and after adult spinal cord lesion. Nature, 367, 170-173 (1994)[PubMed]
- Omoto, S., Ueno, M., Mochio, S. et al.: Corticospinal tract fibers cross the ephrin-B3-negative part of the midline of the spinal cord after brain injury. Neurosci. Res., 69, 187-195 (2011)[PubMed]
- Silver, J., Schwab, M. E. & Popovich, P. G.: Central nervous system regenerative failure: role of oligodendrocytes, astrocytes, and microglia. Cold Spring Harb. Perspect. Biol., 7, a020602 (2014)[PubMed]
- Schwab, M. E. & Caroni, P.: Oligodendrocytes and CNS myelin are nonpermissive substrates for neurite growth and fibroblast spreading in vitro. J. Neurosci., 8, 2381-2393 (1988)[PubMed]
- Lee, J. K., Geoffroy, C. G., Chan, A. F. et al.: Assessing spinal axon regeneration and sprouting in Nogo-, MAG-, and OMgp-deficient mice. Neuron, 66, 663-670 (2010)[PubMed]
- Cafferty, W. B., Duffy, P., Huebner, E. et al.: MAG and OMgp synergize with Nogo-A to restrict axonal growth and neurological recovery after spinal cord trauma. J. Neurosci., 30, 6825-6837 (2010)[PubMed]
- Omoto, S., Ueno, M., Mochio, S. et al.: Genetic deletion of paired immunoglobulin-like receptor B does not promote axonal plasticity or functional recovery after traumatic brain injury. J. Neurosci., 30, 13045-13052 (2010)[PubMed]
- Goldberg, J. L., Klassen, M. P., Hua, Y. et al.: Amacrine-signaled loss of intrinsic axon growth ability by retinal ganglion cells. Science, 296, 1860-1864 (2002)[PubMed]
- Moore, D. L., Blackmore, M. G., Hu, Y. et al.: KLF family members regulate intrinsic axon regeneration ability. Science, 326, 298-301 (2009)[PubMed]
- Blackmore, M. G., Wang, Z., Lerch, J. K. et al.: Kruppel-like Factor 7 engineered for transcriptional activation promotes axon regeneration in the adult corticospinal tract. Proc. Natl. Acad. Sci. USA, 109, 7517-7522 (2012)[PubMed]
- Cai, D., Qiu, J., Cao, Z. et al.: Neuronal cyclic AMP controls the developmental loss in ability of axons to regenerate. J. Neurosci., 21, 4731-4739 (2001)[PubMed]
- Wang, Z., Reynolds, A., Kirry, A. et al.: Overexpression of Sox11 promotes corticospinal tract regeneration after spinal injury while interfering with functional recovery. J. Neurosci., 35, 3139-3145 (2015)[PubMed]
- Liu, K., Lu, Y., Lee, J. K. et al.: PTEN deletion enhances the regenerative ability of adult corticospinal neurons. Nat. Neurosci., 13, 1075-1081 (2010)[PubMed]
- Carmel, J. B., Berrol, L. J., Brus-Ramer, M. et al.: Chronic electrical stimulation of the intact corticospinal system after unilateral injury restores skilled locomotor control and promotes spinal axon outgrowth. J. Neurosci., 30, 10918-10926 (2010)[PubMed]
- Brus-Ramer, M., Carmel, J. B., Chakrabarty, S. et al.: Electrical stimulation of spared corticospinal axons augments connections with ipsilateral spinal motor circuits after injury. J. Neurosci., 27, 13793-13801 (2007)[PubMed]
- Carmichael, S. T. & Chesselet, M. F.: Synchronous neuronal activity is a signal for axonal sprouting after cortical lesions in the adult. J. Neurosci., 22, 6062-6070 (2002)[PubMed]
- Fawcett, J. W.: Recovery from spinal cord injury: regeneration, plasticity and rehabilitation. Brain, 132, 1417-1418 (2009)[PubMed]
- Nakagawa, H., Ueno, M., Itokazu, T. et al.: Bilateral movement training promotes axonal remodeling of the corticospinal tract and recovery of motor function following traumatic brain injury in mice. Cell Death Dis., 4, e534 (2013)[PubMed]
- Taub, E., Crago, J. E. & Uswatte, G.: Constraint-induced movement therapy: a new approach to treatment in physical rehabilitation. Rehabil. Psychol., 43, 152-170 (1998)
- Brown, A. & Weaver, L. C.: The dark side of neuroplasticity. Exp. Neurol., 235, 133-141 (2012)[PubMed]
- Geoffroy, C. G., Lorenzana, A. O., Kwan, J. P. et al.: Effects of PTEN and Nogo codeletion on corticospinal axon sprouting and regeneration in mice. J. Neurosci., 35, 6413-6428 (2015)[PubMed]
- Lu, P., Blesch, A., Graham, L. et al.: Motor axonal regeneration after partial and complete spinal cord transection. J. Neurosci., 32, 8208-8218 (2012)[PubMed]
- Wahl, A. S., Omlor, W., Rubio, J. C. et al.: Asynchronous therapy restores motor control by rewiring of the rat corticospinal tract after stroke. Science, 344, 1250-1255 (2014)[PubMed]
- Takeoka, A., Vollenweider, I., Courtine, G. et al.: Muscle spindle feedback directs locomotor recovery and circuit reorganization after spinal cord injury. Cell, 159, 1626-1639 (2014)[PubMed]
- Bui, T. V., Stifani, N., Akay, T. et al.: Spinal microcircuits comprising dI3 interneurons are necessary for motor functional recovery following spinal cord transection. Elife, 5, e21715 (2016)[PubMed]
- Bellardita, C., Caggiano, V., Leiras, R. et al.: Spatiotemporal correlation of spinal network dynamics underlying spasms in chronic spinalized mice. Elife, 6, e23011 (2017)[PubMed]
- Hilton, B. J., Anenberg, E., Harrison, T. C. et al.: Re-establishment of cortical motor output maps and spontaneous functional recovery via spared dorsolaterally projecting corticospinal neurons after dorsal column spinal cord injury in adult mice. J. Neurosci., 36, 4080-4092 (2016)[PubMed]
- Siegel, C. S., Fink, K. L., Strittmatter, S. M. et al.: Plasticity of intact rubral projections mediates spontaneous recovery of function after corticospinal tract injury. J. Neurosci., 35, 1443-1457 (2015)[PubMed]
著者プロフィール
略歴:2006年 東京大学大学院農学生命科学研究科 修了,理化学研究所脳科学総合研究センター,千葉大学大学院医学研究院,大阪大学大学院医学系研究科,米国Cincinnati Children's Hospital Medical Center,科学技術振興機構さきがけ研究者を経て,2016年より新潟大学脳研究所 特任教授.
研究テーマ:健常な脳および障害をうけた脳における神経回路の構成とその動作原理.
© 2017 上野 将紀 Licensed under CC 表示 2.1 日本
