海馬体-嗅内皮質における空間認知システム
2015/01/08
北西卓磨・松尾直毅
(大阪大学大学院医学系研究科 分子行動神経科学)
email:北西卓磨,松尾直毅
領域融合レビュー, 4, e001 (2015) DOI: 10.7875/leading.author.4.e001
Takuma Kitanishi & Naoki Matsuo: Spatial representation system in the hippocampal-entorhinal circuit.

内側側頭葉の海馬体-嗅内皮質には,場所細胞,格子細胞,頭部方向細胞が存在する.これらのニューロンは,壁やオブジェクトなど外界のランドマークを基準とした空間選択的な発火を示し,動物の空間認知をささえる“地図”としてはたらくと考えられている.また,外界の変化や連合学習に対する活動パターンの様態から,空間記憶やエピソード記憶にも関与すると考えられる.近年になり,空間選択的な活動パターンを生成する回路機構および細胞機構の解明にむけ多彩な研究アプローチが適用されるようになり研究が加速している.
2014年のノーベル医学生理学賞は,“脳の空間認知に関する細胞の発見”について,John O'Keefe(ジョン・オキーフ)教授,May-Britt Moser(マイブリット・モーザー)教授,Edvard I. Moser(エドヴァルド・モーザー)教授の3氏に授与された.このレビューは,受賞の対象となった場所細胞および格子細胞とはどのような細胞なのか解説する.また,これらの細胞を形成する回路機構や記憶との関連について,最新の知見をまじえ議論する.
“いま自分がどこにいるのか?”という空間認知は,われわれが生きていくために重要かつ基本的な感覚である.空間のなかでの自分の居場所を認識することができなければ,知らない土地を旅することはもちろん,毎日の通勤すらままならない.空間認知能力の低下は認知症の中核症状でもある.空間を正しく認知するために,脳はどのようなしくみを備えているのだろうか.
1971年,O'Keefeらは,空間のなかでの動物の位置に反応して活動するニューロンが存在することを発見した1).この研究が行われた当時,すでに海馬という脳部位が記憶において重要であると考えられていた2).そこで彼らは,自身も開発にたずさわった,行動中の動物の脳から神経活動を計測する当時の先端技術を海馬に適用することにした.海馬に留置した微小電極を介してラットが部屋のなかで自由に活動するときの神経活動を計測したところ,ラットが部屋のなかの特定の場所で特定の方向をむいたときにだけ活動するニューロンが存在することを見い出した.そののち,方向選択性は実は平面的な広がりのある部屋では弱く(ただし,1次元の通路状の環境では顕著な方向選択性がある),動物が特定の場所を通り抜けるときにおもに活動することがわかった.これが場所細胞(place cell)である(図1a).
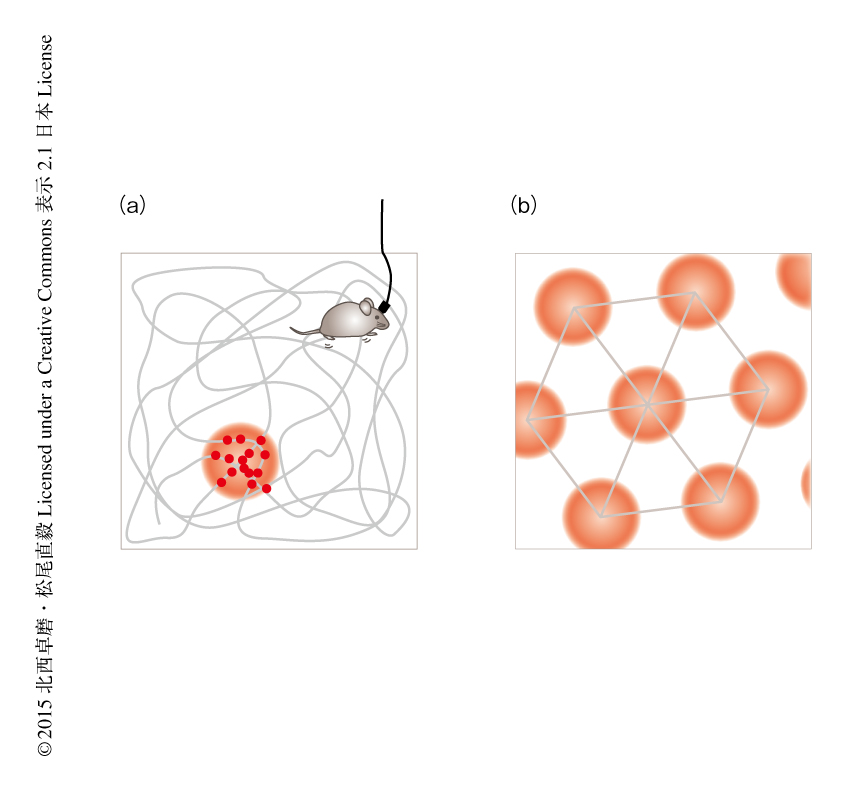
場所細胞が発火活動をしめす場所である場所受容野は,場所細胞ごとにさまざまである.たとえば,ある場所細胞は部屋のある角で活動し,別の場所細胞は部屋の中心で活動するというぐあいである.したがって,多数の場所細胞を集めると,場所受容野が動物の歩きまわる環境全体をおおいつくす.そのため,場所細胞の活動を読み取ることにより逆に,動物がいまどこにいるかを予測できる.実際,同時に計測した数十個の場所細胞から10 cm以下の誤差でラットの現在位置を推定できる3).また,場所細胞のきわだった特徴として,特定の感覚情報に依存しないことがあげられる.たとえば,暗闇にして視覚情報をうばっても場所細胞の活動はもとどおり維持される.嗅覚,触覚,聴覚の情報も不要である4).さらに,動物をカートに乗せて動かしても場所選択的な活動は生じることから,歩くという運動指令の情報も必須ではない5).したがって,場所細胞の活動は特定の感覚情報により一義に決まるのではなく,さまざまな情報を統合した結果として,脳が内的に生成する空間認知を反映するものと考えられる.このような特徴から,場所細胞は認知地図の神経基盤であると提唱された6).
場所細胞の活動パターンは綿密に調べられてきた.さまざまなかたちの環境(円形,多角形,直線,8の字型,E字型,ジグザグ型),プール7),スペースシャトルの無重力空間8) など,多様な環境において場所選択的な活動が確認されている.また,ラットにくわえ,マウス,コウモリ,サルでも場所細胞がみつかっている.なお,コウモリが飛翔するときの場所受容野は球形(高さにより発火率が変わる)であるのに対し9),ラットがらせん階段や壁を登る際の場所受容野は鉛直方向に延びており高さ方向の分解能が低い10).ヒトにおいても,ディスプレイ越しのバーチャルな環境においてではあるが,場所選択的に活動するニューロンの存在が確認されている11).場所細胞は哺乳類に共通の普遍的な空間認知システムを構成すると考えられる.
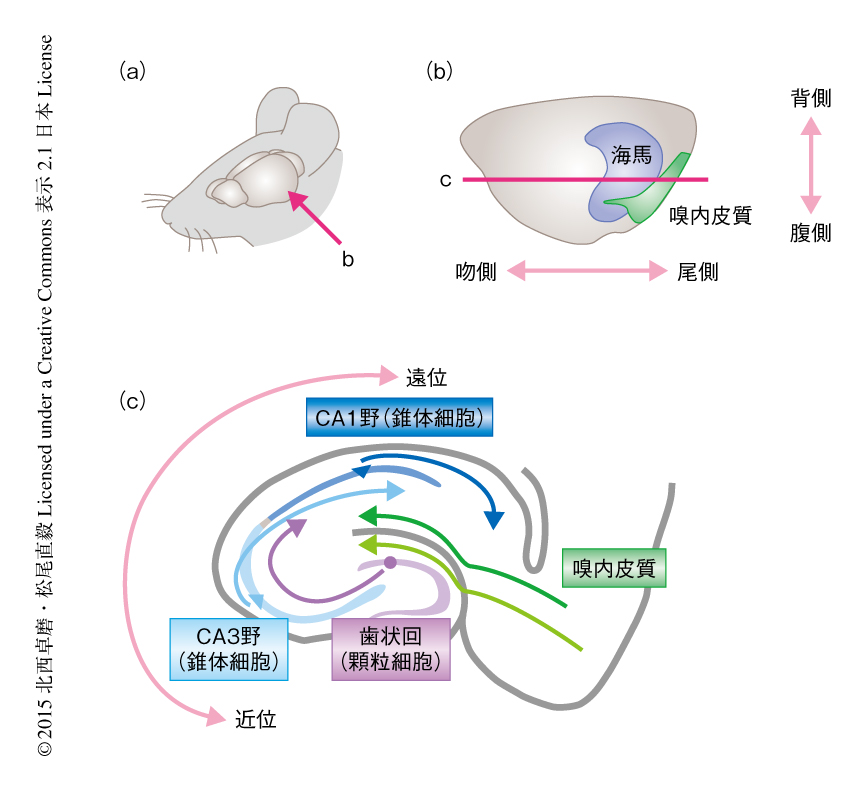
海馬の構造に目をむけて,場所細胞の特徴についてよりくわしくみていこう(図2).海馬のCA1野およびCA3野の錐体細胞と歯状回の顆粒細胞が場所細胞としての活動パターンを示し,介在細胞はいずれの部位においても明確な場所選択性をもたないとされる(ただし,多くの場合,介在細胞か否かという細胞種の分類は実験で得られる電気生理学的な特徴にもとづいており,これが組織学的な分類と厳密に合致するかは不明である).
場所細胞の活動パターンは外部の環境に応じて動的かつ瞬時に再配置することが知られており,この再配置の様態が領域により異なっている.再配置には,場所受容野が変化する包括的な再配置や,場所受容野は変わらず発火率だけが変化する頻度の再配置がある.環境の形状を正方形から円形へとしだいに変形させたときのCA1野における再配置のようすがくわしく調べられ,形状の変化がわずかなら場所受容野はおおよそ一定に保たれる一方,形状の変化が大きくなると場所受容野が一斉に包括的な再配置を示すことが見い出された12).この結果は,場所細胞は,変化あるいは欠落した情報から全体像を補完するパターン完成と,外界のわずかな変化を鋭敏に区別するパターン分離の両方を行うことを示唆している.パターン分離はCA1野の上流である歯状回とCA3野が起点だと考えられている.歯状回では,多くの場所細胞は複数の場所受容野をもち,環境のわずかな差異によりおのおのの場所受容野における発火率が鋭敏に変化する13).CA3野では環境のわずかな変化は頻度の再配置として表現される一方,変化が大きくなるとそれまで発火率の低かったニューロンが新たに場所細胞として活動をはじめたり,逆に,活動していた場所細胞が活動しなくなったりという再配置が生じる13,14).
こうした領域のあいだでの性質の違いにくわえ,同じ領域においても,体軸(背側-腹側,近位-遠位,表層-深層の3軸)にそった位置により場所細胞の性質は異なっている.顕著なのは,海馬の背側-腹側軸にそった場所受容野のサイズの変化である.ラットに長い通路を真っすぐに走らせた場合,背側CA3野の場所受容野のサイズは約1 mだが,腹側にむかうほど大きくなり,もっとも腹側では10 mに達する15).腹側の場所細胞はおおまかな環境を表現し,背側ではその環境のなかでの詳細な位置を示す,という機能の分化があるのかもしれない.
近位-遠位という軸では場所選択性が異なる.内側嗅内皮質から強く投射をうける近位CA1野は,外側嗅内皮質からの投射をうける遠位CA1野よりも高い空間選択性を示す16).内側嗅内皮質にはのちに述べる格子細胞が存在する一方,外側嗅内皮質のニューロンは明確な空間選択性をもたず,この空間あるいは非空間という情報の質的な差異がCA1野の近位-遠位軸に反映されると考えられている.
また,CA1野の錐体細胞層には表層-深層の軸方向に錐体細胞が5~8層ほどならんでいるが,場所細胞となる割合や発火パターンの特性が表層と深層とで異なる17).このように,海馬における領域や領域の内部での位置により,少しずつ異なる情報を表現する場所細胞が分布している.
場所細胞は海馬に依存する空間記憶やエピソード記憶に深く関与すると考えられている.学習にともなう場所細胞の動的な変化の例をみてみよう.
動物が新奇な環境を探索すると,安定な場所細胞の活動が数分から数十分のうちに形成され18),数日ないし数カ月後19,20) に同じ環境にきても同じ活動パターンが再現されうる.こうした場所細胞の活動パターンは,環境そのものについての空間記憶の形成および維持を反映するようにみえる.
こうした環境自体の学習にくわえ,空間情報とほかの情報とを連合する学習においても場所細胞の活動は変化する.たとえば,環境のなかで電気ショックをあたえる恐怖条件付け学習により場所受容野が再配置する21).また,水迷路学習を行うとゴール地点に場所受容野が集積し7),特定の場所に隠されたえさを集める課題では,えさのある場所へと場所受容野が再配置する22).場所受容野は動物にとり重要な意味をもつ場所に集中的に配置されるのかもしれない.
このような空間情報とほかの情報との連合はどの脳部位において行われるのだろうか.じつは,海馬にはさまざまな感覚情報も入力しており23,24),ものの位置や匂い25),時間26) により発火頻度の変化するニューロンが存在する.さらに,これらのニューロンはしばしば場所選択性を兼ね備える26).こうした複数の情報を連合した神経活動が,学習による場所細胞の再配置の基盤なのかもしれない.
空間情報の処理は海馬だけで行われるわけではない.2005年,May-Britt MoserとEdvard I. Moserらのグループは,内側嗅内皮質に格子細胞が存在することを発見した(図1b).2人はO'Keefeの研究室で学んだのち,当初は場所細胞の研究を行っていたが,しだいに海馬への入出力を担う嗅内皮質に目をむけるようになった.嗅内皮質と海馬は背側どうしおよび腹側どうしが密に連絡している.また,さきに述べたように,海馬では背側の場所細胞ほど鋭い場所選択性を示す.この解剖学的な関係から,嗅内皮質においても背側ほど空間情報の処理に重要だと予想し,この部位から神経活動を計測した.そして実際に,場所選択的な発火活動を見い出すことに成功した27).嗅内皮質のニューロンは,場所細胞とは異なり,環境のなかで多数の受容野をもっていた.この受容野の配列をみきわめるため,より大きな環境を用いて記録を行ったところ受容野が正三角形をしきつめた格子の頂点に規則的に配置することを発見し,格子細胞(grid cell)と名づけた28).腹側ほど格子細胞の受容野の間隔が拡大することもわかった.のちに,格子細胞は前海馬支脚および傍海馬支脚においてもみつかっている29).格子細胞の受容野のあいだの距離は環境によらずおおよそ一定である.また,格子細胞は場所細胞とは異なり,どの環境でも活動しつづけ経験による影響をさほどうけない(ただし,論文30,31) も参照のこと).これらの性質から,彼らは,格子細胞は普遍的な距離の指標であると提唱している.
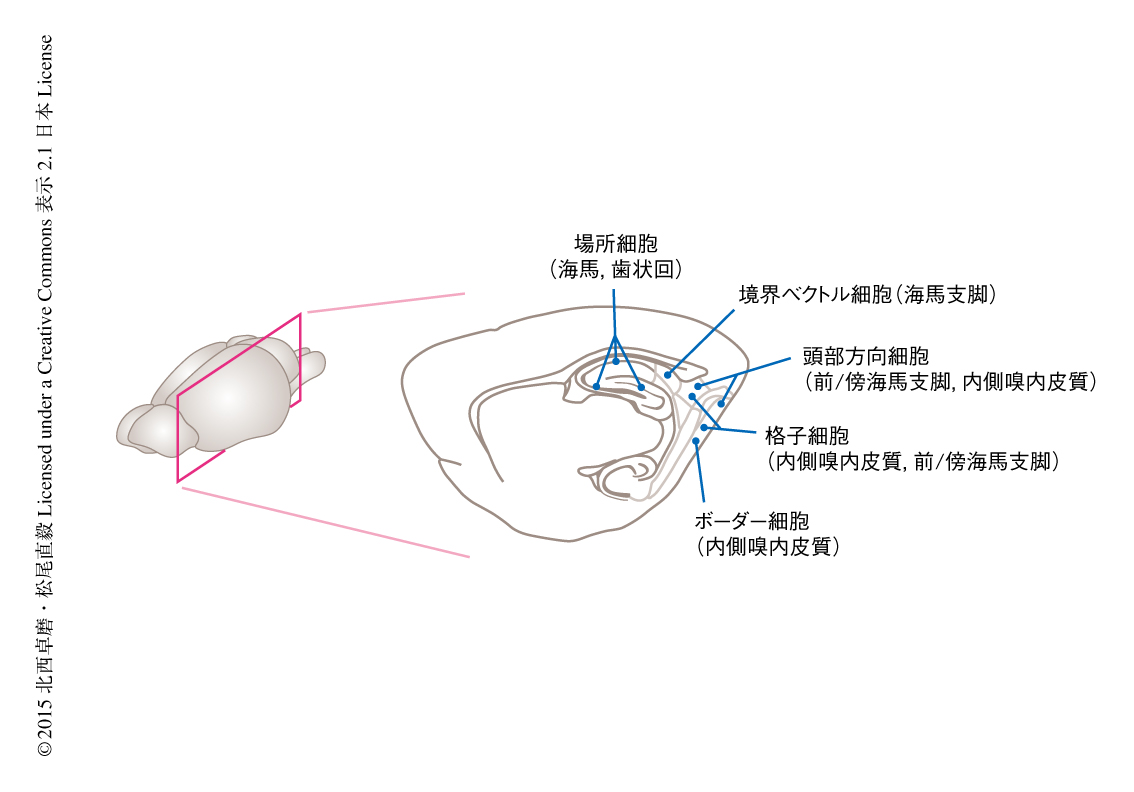
海馬体-嗅内皮質には,ほかにも空間選択的なニューロンが存在する(図3).前海馬支脚において,動物の頭の向きに反応して発火する頭部方向細胞(head direction cell)が発見されている32,33).そののち,頭部方向細胞は内側嗅内皮質,背側被蓋核,外側乳頭体核,視床前核,膨大後部皮質,傍海馬支脚にも存在すると報告された34).また,海馬支脚には,動物から一定の方向かつ一定の距離に壁が存在するときに活動する境界ベクトル細胞(boundary vector cell)がみつかっている35).内側嗅内皮質には,格子細胞と頭部方向細胞の両方の性質をあわせもつコンジャンクティブ細胞(conjunctive cell)や36),壁際などの環境の端にいるときに活動するボーダー細胞(border cell)も存在する37).また,外側嗅内皮質のニューロンは,動物のいる場所ではなく,部屋のなかに置かれたオブジェクトの場所に反応する38).
こうしたさまざまな空間情報はどのように統合されて活用されるのだろうか? そのひとつの仮説は,経路積分とランドマークによる補正とを組み合わせた空間ナビゲーションである39).現在地から別の地点へと真っすぐ移動することを想定してみよう.まず,場所細胞が現在地を示し,頭部方向細胞が移動方向をさし示す.格子細胞の周期的な発火活動を移動距離の指標として用いると到着地点の座標が計算できる.これを小刻みにくり返せば,任意の曲線にそった経路積分によるナビゲーションとなる.ただし,経路積分は内的な空間情報を用いたナビゲーションであり,しだいに実環境との誤差が蓄積する.境界ベクトル細胞やボーダー細胞は壁やオブジェクトなど外界のランドマークからの距離を測り直し,経路積分の誤差を修正するために役だっているのかもしれない.
空間選択的なニューロンは海馬体-嗅内皮質に密集しており,これらの活動にはなんらかの相互作用があると想定される.しかし,こうした相互作用や空間選択的な活動を形成するための回路機構には,まだ不透明な点が多い.
格子細胞が報告された当初,格子細胞からの入力をうけて場所細胞が形成されるというモデルが提唱された40).格子細胞は海馬に投射する表層の嗅内皮質に多く存在し36),実際に,海馬へと投射する電気生理学的な証拠があることからも13,41),この仮説はいっけん合理的に思えた.しかし,近年の研究からは,このモデルに疑問を投げかける知見が集まっている.海馬および嗅内皮質に投射する内側中隔核を抑制すると,格子細胞の活動の周期性が失われるにもかかわらず場所細胞の活動には影響せず42),新奇の環境においては新しい場所細胞も形成されることが報告された43).さらに,背側から腹側にいたる内側嗅内皮質を完全に破壊した場合でも,場所細胞の活動は部分的にしか障害されなかった44).これに対し,逆に,海馬の活動を抑制すると格子細胞の活動が失われる45).嗅内皮質と海馬は,上流と下流という一方向性の階層関係ではなく,相互作用により安定な空間表現をつくる相補的なシステムなのかもしれない.そもそも,場所細胞や格子細胞のもつ空間情報はどの脳領域からもたらされるのだろうか? 頭部方向細胞の加算により格子細胞が形成されるという説や,境界ベクトル細胞の加算により場所受容野が形成されるとする説もあるが,今後の研究が待たれる.
上流からの入力パターン自体にくわえて,その入力がシナプスを介してニューロンに伝達される効率も空間選択的な活動の確立においては重要であろう.シナプス可塑性による伝達強度の変化は,記憶の基盤となる細胞機構であるとともに,場所細胞の形成および維持にも関与することが示唆されている.シナプス可塑性を抑制するNMDA受容体阻害薬を腹腔内に投与すると,慣れた環境における場所細胞の活動は影響をうけないものの,新奇の環境において形成された場所細胞の活動の長期安定性が失われる46).また,領域選択的にNMDA受容体をノックアウトした場合には,空間記憶の障害とともに場所細胞の発火パターンがくずれる47).シナプス可塑性に関与するタンパク質として,ほかにはCaMKII 48),CREB 49),PKA 50),GluR1 51) が場所細胞の活動に影響すると報告されている.近年の覚醒下におけるパッチクランプ法52) やCa2+イメージング法53) を用いた研究は,場所細胞の樹状突起における非線型の過程が空間選択性の確立に重要であることを強調しており,樹状突起-シナプス部位における演算もみのがせない.
空間情報処理の動的な側面をとらえるため,生理的な条件における領域間の情報伝達を探求するアプローチもある.こうしたアプローチの例として,ここではガンマ波に着目した研究について紹介する.ガンマ波は海馬体-嗅内皮質を含むさまざまな脳領域で観察される周波数30~90 Hzの脳波である.ニューロンの活動パターンはガンマ波の影響をうけ,しばしば,ガンマ波の特定の位相で発火するようタイミングが固定される.この現象を位相固定という.ニューロンが集団で特定のガンマ波に位相固定すると,ミリ秒の精度で同期した発火となりシナプス後細胞に活動が伝達されやすい.そのため,ガンマ波への位相固定は領域間の効率的な情報伝達に貢献すると考えられている.
海馬体-嗅内皮質では,遅いガンマ波と速いガンマ波の2種類の周波数帯のガンマ波が観察される.CA3野→CA1野という経路では遅いガンマ波が,嗅内皮質→CA1野という経路では速いガンマ波が,それぞれ同期して生じ,CA1野にはおのおののガンマ波に位相固定する場所細胞の存在することが報告された54).また,この2つの経路におけるガンマ波の同期性や位相固定の強さは行動に応じて変化する55-58).そのため,上流のCA3野および嗅内皮質はおのおののガンマ波の帯域の神経活動を介して,CA1野の場所細胞の活動をダイナミックに制御するものと考えられる.
こうした知見から,ガンマ波への位相固定は場所細胞の活動パターンに密接に関連すると考えられた.しかし,位相固定を制御する細胞機構や,位相固定と場所細胞の活動との具体的な関係は不明だった.そこで筆者のひとり(北西)は,行動中のラットのCA1野からマルチユニット記録を行い,新奇の経験に際した位相固定の様態と,その細胞機構としてシナプス可塑性の関与について調べた.
一般的な薬理学的な手法あるいは遺伝学的な手法によるシナプス可塑性の操作では,脳機能や行動を全身性に障害することが多い.そのため,こうした操作により生じた神経活動の変化は,シナプス可塑性の操作により直接的に生じたのか,あるいは,全身性の変化により間接的に生じたのかを区別できない.この問題を解決するため,ウイルスベクターによる局所的な遺伝子改変と,その部位から正確にマルチユニット記録を行う手法を新たに確立した(図4a).この手法により,正常な脳環境において,CA1野の狭い領域におけるシナプス可塑性とその領域の神経活動パターンとの因果関係を調べることが可能になった.
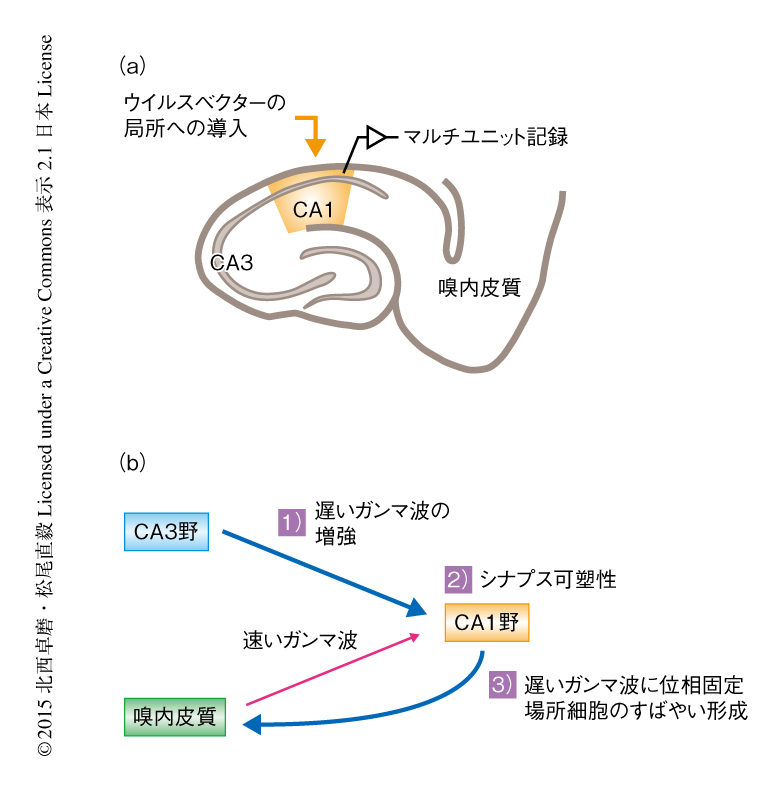
動物が新奇の環境を探索するときには遅いガンマ波が一時的に増強し,このガンマ波へのCA1野の錐体細胞の位相固定が強まることがわかった.さらに,位相固定の強いニューロンほど,空間選択性の高い場所細胞の活動を形成した.一方で,CA1野の錐体細胞のシナプス可塑性を阻害すると,遅いガンマ波への位相固定が減弱し,場所細胞の形成も遅れることが明らかになった.こうした結果と,ガンマ波の領域間の同期を考慮すると,空間学習の際の海馬回路の挙動が推定された(図4b).シナプス可塑性は,細胞レベルにおいては,個々のニューロンの時間的あるいは空間的な発火パターンの形成に寄与するとともに,回路レベルにおいては,記憶形成の際にCA3野→CA1野→嗅内皮質という経路の情報伝達を選択的に増強する役割をもつと考えられる.こうした知見を積み重ねることにより,場所細胞と空間記憶との関連をより明確にできるはずである.
場所細胞と格子細胞を中心に,海馬体-嗅内皮質における空間認知システムについて解説した.これまでの研究により,場所細胞や格子細胞それ自体の活動パターンについては全体像がみえてきた.今後は,こうした活動がどのような“入力”により生成され,できあがった空間選択的な“出力”が高次機能にどう活用されるかを理解することが課題になるだろう.研究手法についても,このレビューにおいて紹介した局所的な遺伝子改変法にくわえ,行動中に活動したニューロンを選択的に操作する特殊な遺伝子改変マウス59,60) や,光遺伝学的な手法,薬理遺伝学な手法など,より選択性および自由度の高い方法を適用することが可能になりつつある.こうした方法を活用し,あるいは,必要に応じて開発し,生きた脳におけるニューロンの入出力演算を探求していくことが重要である.
略歴:2009年 東京大学大学院薬学系研究科 修了,同年 ノルウェーNorwegian University of Science and Technology博士研究員,2013年 京都大学生命科学系キャリアパス形成ユニット 研究員を経て,2014年より大阪大学大学院医学系研究科 研究員.
研究テーマ:海馬および嗅内皮質における情報処理の機構.脳に流れる情報を手に取るように見たい.
松尾 直毅(Naoki Matsuo)
大阪大学大学院医学系研究科 独立准教授.
© 2015 北西卓磨・松尾直毅 Licensed under CC 表示 2.1 日本
(大阪大学大学院医学系研究科 分子行動神経科学)
email:北西卓磨,松尾直毅
領域融合レビュー, 4, e001 (2015) DOI: 10.7875/leading.author.4.e001
Takuma Kitanishi & Naoki Matsuo: Spatial representation system in the hippocampal-entorhinal circuit.
要 約
内側側頭葉の海馬体-嗅内皮質には,場所細胞,格子細胞,頭部方向細胞が存在する.これらのニューロンは,壁やオブジェクトなど外界のランドマークを基準とした空間選択的な発火を示し,動物の空間認知をささえる“地図”としてはたらくと考えられている.また,外界の変化や連合学習に対する活動パターンの様態から,空間記憶やエピソード記憶にも関与すると考えられる.近年になり,空間選択的な活動パターンを生成する回路機構および細胞機構の解明にむけ多彩な研究アプローチが適用されるようになり研究が加速している.
はじめに
2014年のノーベル医学生理学賞は,“脳の空間認知に関する細胞の発見”について,John O'Keefe(ジョン・オキーフ)教授,May-Britt Moser(マイブリット・モーザー)教授,Edvard I. Moser(エドヴァルド・モーザー)教授の3氏に授与された.このレビューは,受賞の対象となった場所細胞および格子細胞とはどのような細胞なのか解説する.また,これらの細胞を形成する回路機構や記憶との関連について,最新の知見をまじえ議論する.
1.場所細胞の発見
“いま自分がどこにいるのか?”という空間認知は,われわれが生きていくために重要かつ基本的な感覚である.空間のなかでの自分の居場所を認識することができなければ,知らない土地を旅することはもちろん,毎日の通勤すらままならない.空間認知能力の低下は認知症の中核症状でもある.空間を正しく認知するために,脳はどのようなしくみを備えているのだろうか.
1971年,O'Keefeらは,空間のなかでの動物の位置に反応して活動するニューロンが存在することを発見した1).この研究が行われた当時,すでに海馬という脳部位が記憶において重要であると考えられていた2).そこで彼らは,自身も開発にたずさわった,行動中の動物の脳から神経活動を計測する当時の先端技術を海馬に適用することにした.海馬に留置した微小電極を介してラットが部屋のなかで自由に活動するときの神経活動を計測したところ,ラットが部屋のなかの特定の場所で特定の方向をむいたときにだけ活動するニューロンが存在することを見い出した.そののち,方向選択性は実は平面的な広がりのある部屋では弱く(ただし,1次元の通路状の環境では顕著な方向選択性がある),動物が特定の場所を通り抜けるときにおもに活動することがわかった.これが場所細胞(place cell)である(図1a).
場所細胞が発火活動をしめす場所である場所受容野は,場所細胞ごとにさまざまである.たとえば,ある場所細胞は部屋のある角で活動し,別の場所細胞は部屋の中心で活動するというぐあいである.したがって,多数の場所細胞を集めると,場所受容野が動物の歩きまわる環境全体をおおいつくす.そのため,場所細胞の活動を読み取ることにより逆に,動物がいまどこにいるかを予測できる.実際,同時に計測した数十個の場所細胞から10 cm以下の誤差でラットの現在位置を推定できる3).また,場所細胞のきわだった特徴として,特定の感覚情報に依存しないことがあげられる.たとえば,暗闇にして視覚情報をうばっても場所細胞の活動はもとどおり維持される.嗅覚,触覚,聴覚の情報も不要である4).さらに,動物をカートに乗せて動かしても場所選択的な活動は生じることから,歩くという運動指令の情報も必須ではない5).したがって,場所細胞の活動は特定の感覚情報により一義に決まるのではなく,さまざまな情報を統合した結果として,脳が内的に生成する空間認知を反映するものと考えられる.このような特徴から,場所細胞は認知地図の神経基盤であると提唱された6).
場所細胞の活動パターンは綿密に調べられてきた.さまざまなかたちの環境(円形,多角形,直線,8の字型,E字型,ジグザグ型),プール7),スペースシャトルの無重力空間8) など,多様な環境において場所選択的な活動が確認されている.また,ラットにくわえ,マウス,コウモリ,サルでも場所細胞がみつかっている.なお,コウモリが飛翔するときの場所受容野は球形(高さにより発火率が変わる)であるのに対し9),ラットがらせん階段や壁を登る際の場所受容野は鉛直方向に延びており高さ方向の分解能が低い10).ヒトにおいても,ディスプレイ越しのバーチャルな環境においてではあるが,場所選択的に活動するニューロンの存在が確認されている11).場所細胞は哺乳類に共通の普遍的な空間認知システムを構成すると考えられる.
2.場所細胞の解剖学
海馬の構造に目をむけて,場所細胞の特徴についてよりくわしくみていこう(図2).海馬のCA1野およびCA3野の錐体細胞と歯状回の顆粒細胞が場所細胞としての活動パターンを示し,介在細胞はいずれの部位においても明確な場所選択性をもたないとされる(ただし,多くの場合,介在細胞か否かという細胞種の分類は実験で得られる電気生理学的な特徴にもとづいており,これが組織学的な分類と厳密に合致するかは不明である).
場所細胞の活動パターンは外部の環境に応じて動的かつ瞬時に再配置することが知られており,この再配置の様態が領域により異なっている.再配置には,場所受容野が変化する包括的な再配置や,場所受容野は変わらず発火率だけが変化する頻度の再配置がある.環境の形状を正方形から円形へとしだいに変形させたときのCA1野における再配置のようすがくわしく調べられ,形状の変化がわずかなら場所受容野はおおよそ一定に保たれる一方,形状の変化が大きくなると場所受容野が一斉に包括的な再配置を示すことが見い出された12).この結果は,場所細胞は,変化あるいは欠落した情報から全体像を補完するパターン完成と,外界のわずかな変化を鋭敏に区別するパターン分離の両方を行うことを示唆している.パターン分離はCA1野の上流である歯状回とCA3野が起点だと考えられている.歯状回では,多くの場所細胞は複数の場所受容野をもち,環境のわずかな差異によりおのおのの場所受容野における発火率が鋭敏に変化する13).CA3野では環境のわずかな変化は頻度の再配置として表現される一方,変化が大きくなるとそれまで発火率の低かったニューロンが新たに場所細胞として活動をはじめたり,逆に,活動していた場所細胞が活動しなくなったりという再配置が生じる13,14).
こうした領域のあいだでの性質の違いにくわえ,同じ領域においても,体軸(背側-腹側,近位-遠位,表層-深層の3軸)にそった位置により場所細胞の性質は異なっている.顕著なのは,海馬の背側-腹側軸にそった場所受容野のサイズの変化である.ラットに長い通路を真っすぐに走らせた場合,背側CA3野の場所受容野のサイズは約1 mだが,腹側にむかうほど大きくなり,もっとも腹側では10 mに達する15).腹側の場所細胞はおおまかな環境を表現し,背側ではその環境のなかでの詳細な位置を示す,という機能の分化があるのかもしれない.
近位-遠位という軸では場所選択性が異なる.内側嗅内皮質から強く投射をうける近位CA1野は,外側嗅内皮質からの投射をうける遠位CA1野よりも高い空間選択性を示す16).内側嗅内皮質にはのちに述べる格子細胞が存在する一方,外側嗅内皮質のニューロンは明確な空間選択性をもたず,この空間あるいは非空間という情報の質的な差異がCA1野の近位-遠位軸に反映されると考えられている.
また,CA1野の錐体細胞層には表層-深層の軸方向に錐体細胞が5~8層ほどならんでいるが,場所細胞となる割合や発火パターンの特性が表層と深層とで異なる17).このように,海馬における領域や領域の内部での位置により,少しずつ異なる情報を表現する場所細胞が分布している.
3.場所細胞と記憶
場所細胞は海馬に依存する空間記憶やエピソード記憶に深く関与すると考えられている.学習にともなう場所細胞の動的な変化の例をみてみよう.
動物が新奇な環境を探索すると,安定な場所細胞の活動が数分から数十分のうちに形成され18),数日ないし数カ月後19,20) に同じ環境にきても同じ活動パターンが再現されうる.こうした場所細胞の活動パターンは,環境そのものについての空間記憶の形成および維持を反映するようにみえる.
こうした環境自体の学習にくわえ,空間情報とほかの情報とを連合する学習においても場所細胞の活動は変化する.たとえば,環境のなかで電気ショックをあたえる恐怖条件付け学習により場所受容野が再配置する21).また,水迷路学習を行うとゴール地点に場所受容野が集積し7),特定の場所に隠されたえさを集める課題では,えさのある場所へと場所受容野が再配置する22).場所受容野は動物にとり重要な意味をもつ場所に集中的に配置されるのかもしれない.
このような空間情報とほかの情報との連合はどの脳部位において行われるのだろうか.じつは,海馬にはさまざまな感覚情報も入力しており23,24),ものの位置や匂い25),時間26) により発火頻度の変化するニューロンが存在する.さらに,これらのニューロンはしばしば場所選択性を兼ね備える26).こうした複数の情報を連合した神経活動が,学習による場所細胞の再配置の基盤なのかもしれない.
4.格子細胞
空間情報の処理は海馬だけで行われるわけではない.2005年,May-Britt MoserとEdvard I. Moserらのグループは,内側嗅内皮質に格子細胞が存在することを発見した(図1b).2人はO'Keefeの研究室で学んだのち,当初は場所細胞の研究を行っていたが,しだいに海馬への入出力を担う嗅内皮質に目をむけるようになった.嗅内皮質と海馬は背側どうしおよび腹側どうしが密に連絡している.また,さきに述べたように,海馬では背側の場所細胞ほど鋭い場所選択性を示す.この解剖学的な関係から,嗅内皮質においても背側ほど空間情報の処理に重要だと予想し,この部位から神経活動を計測した.そして実際に,場所選択的な発火活動を見い出すことに成功した27).嗅内皮質のニューロンは,場所細胞とは異なり,環境のなかで多数の受容野をもっていた.この受容野の配列をみきわめるため,より大きな環境を用いて記録を行ったところ受容野が正三角形をしきつめた格子の頂点に規則的に配置することを発見し,格子細胞(grid cell)と名づけた28).腹側ほど格子細胞の受容野の間隔が拡大することもわかった.のちに,格子細胞は前海馬支脚および傍海馬支脚においてもみつかっている29).格子細胞の受容野のあいだの距離は環境によらずおおよそ一定である.また,格子細胞は場所細胞とは異なり,どの環境でも活動しつづけ経験による影響をさほどうけない(ただし,論文30,31) も参照のこと).これらの性質から,彼らは,格子細胞は普遍的な距離の指標であると提唱している.
5.そのほかの空間選択的なニューロン
海馬体-嗅内皮質には,ほかにも空間選択的なニューロンが存在する(図3).前海馬支脚において,動物の頭の向きに反応して発火する頭部方向細胞(head direction cell)が発見されている32,33).そののち,頭部方向細胞は内側嗅内皮質,背側被蓋核,外側乳頭体核,視床前核,膨大後部皮質,傍海馬支脚にも存在すると報告された34).また,海馬支脚には,動物から一定の方向かつ一定の距離に壁が存在するときに活動する境界ベクトル細胞(boundary vector cell)がみつかっている35).内側嗅内皮質には,格子細胞と頭部方向細胞の両方の性質をあわせもつコンジャンクティブ細胞(conjunctive cell)や36),壁際などの環境の端にいるときに活動するボーダー細胞(border cell)も存在する37).また,外側嗅内皮質のニューロンは,動物のいる場所ではなく,部屋のなかに置かれたオブジェクトの場所に反応する38).
こうしたさまざまな空間情報はどのように統合されて活用されるのだろうか? そのひとつの仮説は,経路積分とランドマークによる補正とを組み合わせた空間ナビゲーションである39).現在地から別の地点へと真っすぐ移動することを想定してみよう.まず,場所細胞が現在地を示し,頭部方向細胞が移動方向をさし示す.格子細胞の周期的な発火活動を移動距離の指標として用いると到着地点の座標が計算できる.これを小刻みにくり返せば,任意の曲線にそった経路積分によるナビゲーションとなる.ただし,経路積分は内的な空間情報を用いたナビゲーションであり,しだいに実環境との誤差が蓄積する.境界ベクトル細胞やボーダー細胞は壁やオブジェクトなど外界のランドマークからの距離を測り直し,経路積分の誤差を修正するために役だっているのかもしれない.
6.空間選択的な神経活動の回路機構および細胞機構
空間選択的なニューロンは海馬体-嗅内皮質に密集しており,これらの活動にはなんらかの相互作用があると想定される.しかし,こうした相互作用や空間選択的な活動を形成するための回路機構には,まだ不透明な点が多い.
格子細胞が報告された当初,格子細胞からの入力をうけて場所細胞が形成されるというモデルが提唱された40).格子細胞は海馬に投射する表層の嗅内皮質に多く存在し36),実際に,海馬へと投射する電気生理学的な証拠があることからも13,41),この仮説はいっけん合理的に思えた.しかし,近年の研究からは,このモデルに疑問を投げかける知見が集まっている.海馬および嗅内皮質に投射する内側中隔核を抑制すると,格子細胞の活動の周期性が失われるにもかかわらず場所細胞の活動には影響せず42),新奇の環境においては新しい場所細胞も形成されることが報告された43).さらに,背側から腹側にいたる内側嗅内皮質を完全に破壊した場合でも,場所細胞の活動は部分的にしか障害されなかった44).これに対し,逆に,海馬の活動を抑制すると格子細胞の活動が失われる45).嗅内皮質と海馬は,上流と下流という一方向性の階層関係ではなく,相互作用により安定な空間表現をつくる相補的なシステムなのかもしれない.そもそも,場所細胞や格子細胞のもつ空間情報はどの脳領域からもたらされるのだろうか? 頭部方向細胞の加算により格子細胞が形成されるという説や,境界ベクトル細胞の加算により場所受容野が形成されるとする説もあるが,今後の研究が待たれる.
上流からの入力パターン自体にくわえて,その入力がシナプスを介してニューロンに伝達される効率も空間選択的な活動の確立においては重要であろう.シナプス可塑性による伝達強度の変化は,記憶の基盤となる細胞機構であるとともに,場所細胞の形成および維持にも関与することが示唆されている.シナプス可塑性を抑制するNMDA受容体阻害薬を腹腔内に投与すると,慣れた環境における場所細胞の活動は影響をうけないものの,新奇の環境において形成された場所細胞の活動の長期安定性が失われる46).また,領域選択的にNMDA受容体をノックアウトした場合には,空間記憶の障害とともに場所細胞の発火パターンがくずれる47).シナプス可塑性に関与するタンパク質として,ほかにはCaMKII 48),CREB 49),PKA 50),GluR1 51) が場所細胞の活動に影響すると報告されている.近年の覚醒下におけるパッチクランプ法52) やCa2+イメージング法53) を用いた研究は,場所細胞の樹状突起における非線型の過程が空間選択性の確立に重要であることを強調しており,樹状突起-シナプス部位における演算もみのがせない.
7.ガンマ波に着目した回路機構の解明
空間情報処理の動的な側面をとらえるため,生理的な条件における領域間の情報伝達を探求するアプローチもある.こうしたアプローチの例として,ここではガンマ波に着目した研究について紹介する.ガンマ波は海馬体-嗅内皮質を含むさまざまな脳領域で観察される周波数30~90 Hzの脳波である.ニューロンの活動パターンはガンマ波の影響をうけ,しばしば,ガンマ波の特定の位相で発火するようタイミングが固定される.この現象を位相固定という.ニューロンが集団で特定のガンマ波に位相固定すると,ミリ秒の精度で同期した発火となりシナプス後細胞に活動が伝達されやすい.そのため,ガンマ波への位相固定は領域間の効率的な情報伝達に貢献すると考えられている.
海馬体-嗅内皮質では,遅いガンマ波と速いガンマ波の2種類の周波数帯のガンマ波が観察される.CA3野→CA1野という経路では遅いガンマ波が,嗅内皮質→CA1野という経路では速いガンマ波が,それぞれ同期して生じ,CA1野にはおのおののガンマ波に位相固定する場所細胞の存在することが報告された54).また,この2つの経路におけるガンマ波の同期性や位相固定の強さは行動に応じて変化する55-58).そのため,上流のCA3野および嗅内皮質はおのおののガンマ波の帯域の神経活動を介して,CA1野の場所細胞の活動をダイナミックに制御するものと考えられる.
こうした知見から,ガンマ波への位相固定は場所細胞の活動パターンに密接に関連すると考えられた.しかし,位相固定を制御する細胞機構や,位相固定と場所細胞の活動との具体的な関係は不明だった.そこで筆者のひとり(北西)は,行動中のラットのCA1野からマルチユニット記録を行い,新奇の経験に際した位相固定の様態と,その細胞機構としてシナプス可塑性の関与について調べた.
一般的な薬理学的な手法あるいは遺伝学的な手法によるシナプス可塑性の操作では,脳機能や行動を全身性に障害することが多い.そのため,こうした操作により生じた神経活動の変化は,シナプス可塑性の操作により直接的に生じたのか,あるいは,全身性の変化により間接的に生じたのかを区別できない.この問題を解決するため,ウイルスベクターによる局所的な遺伝子改変と,その部位から正確にマルチユニット記録を行う手法を新たに確立した(図4a).この手法により,正常な脳環境において,CA1野の狭い領域におけるシナプス可塑性とその領域の神経活動パターンとの因果関係を調べることが可能になった.
動物が新奇の環境を探索するときには遅いガンマ波が一時的に増強し,このガンマ波へのCA1野の錐体細胞の位相固定が強まることがわかった.さらに,位相固定の強いニューロンほど,空間選択性の高い場所細胞の活動を形成した.一方で,CA1野の錐体細胞のシナプス可塑性を阻害すると,遅いガンマ波への位相固定が減弱し,場所細胞の形成も遅れることが明らかになった.こうした結果と,ガンマ波の領域間の同期を考慮すると,空間学習の際の海馬回路の挙動が推定された(図4b).シナプス可塑性は,細胞レベルにおいては,個々のニューロンの時間的あるいは空間的な発火パターンの形成に寄与するとともに,回路レベルにおいては,記憶形成の際にCA3野→CA1野→嗅内皮質という経路の情報伝達を選択的に増強する役割をもつと考えられる.こうした知見を積み重ねることにより,場所細胞と空間記憶との関連をより明確にできるはずである.
おわりに
場所細胞と格子細胞を中心に,海馬体-嗅内皮質における空間認知システムについて解説した.これまでの研究により,場所細胞や格子細胞それ自体の活動パターンについては全体像がみえてきた.今後は,こうした活動がどのような“入力”により生成され,できあがった空間選択的な“出力”が高次機能にどう活用されるかを理解することが課題になるだろう.研究手法についても,このレビューにおいて紹介した局所的な遺伝子改変法にくわえ,行動中に活動したニューロンを選択的に操作する特殊な遺伝子改変マウス59,60) や,光遺伝学的な手法,薬理遺伝学な手法など,より選択性および自由度の高い方法を適用することが可能になりつつある.こうした方法を活用し,あるいは,必要に応じて開発し,生きた脳におけるニューロンの入出力演算を探求していくことが重要である.
文 献
- O'Keefe, J. & Dostrovsky, J.: The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res., 34, 171-175 (1971)[PubMed]
- Scoville, W. B. & Milner, B.: Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry, 20, 11-21 (1957)[PubMed]
- Wilson, M. A. & McNaughton, B. L.: Dynamics of the hippocampal ensemble code for space. Science, 261, 1055-1058 (1993)[PubMed]
- O'Keefe, J.: Place units in the hippocampus of the freely moving rat. Exp. Neurol., 51, 78-109 (1976)[PubMed]
- Terrazas, A., Krause, M., Lipa, P. et al.: Self-motion and the hippocampal spatial metric. J. Neurosci., 25, 8085-8096 (2005)[PubMed]
- O'Keefe, J. & Nadel, L.: The Hippocampus as a Cognitive Map. Oxford University Press, Oxford (1978)
- Hollup, S. A., Molden, S., Donnett, J. G. et al.: Accumulation of hippocampal place fields at the goal location in an annular watermaze task. J. Neurosci., 21, 1635-1644 (2001)[PubMed]
- Knierim, J. J., McNaughton, B. L. & Poe, G. R.: Three-dimensional spatial selectivity of hippocampal neurons during space flight. Nat. Neurosci., 3, 209-210 (2000)[PubMed]
- Yartsev, M. M. & Ulanovsky, N.: Representation of three-dimensional space in the hippocampus of flying bats. Science, 340, 367-372 (2013)[PubMed]
- Hayman, R., Verriotis, M. A., Jovalekic, A. et al.: Anisotropic encoding of three-dimensional space by place cells and grid cells. Nat. Neurosci., 14, 1182-1188 (2011)[PubMed]
- Ekstrom, A. D., Kahana, M. J., Caplan, J. B. et al.: Cellular networks underlying human spatial navigation. Nature, 425, 184-188 (2003)[PubMed]
- Wills, T. J., Lever, C., Cacucci, F. et al.: Attractor dynamics in the hippocampal representation of the local environment. Science, 308, 873-876 (2005)[PubMed]
- Leutgeb, J. K., Leutgeb, S., Moser, M. B. et al.: Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science, 315, 961-966 (2007)[PubMed]
- Leutgeb, S., Leutgeb, J. K., Barnes, C. A. et al.: Independent codes for spatial and episodic memory in hippocampal neuronal ensembles. Science, 309, 619-623 (2005)[PubMed]
- Kjelstrup, K. B., Solstad, T., Brun, V. H. et al.: Finite scale of spatial representation in the hippocampus. Science, 321, 140-143 (2008)[PubMed]
- Henriksen, E. J., Colgin, L. L., Barnes, C. A. et al.: Spatial representation along the proximodistal axis of CA1. Neuron, 68, 127-137 (2010)[PubMed]
- Mizuseki, K., Diba, K., Pastalkova, E. et al.: Hippocampal CA1 pyramidal cells form functionally distinct sublayers. Nat. Neurosci., 14, 1174-1181 (2011)[PubMed]
- Frank, L. M., Stanley, G. B. & Brown, E. N.: Hippocampal plasticity across multiple days of exposure to novel environments. J. Neurosci., 24, 7681-7689 (2004)[PubMed]
- Thompson, L. T. & Best, P. J.: Long-term stability of the place-field activity of single units recorded from the dorsal hippocampus of freely behaving rats. Brain Res., 509, 299-308 (1990)[PubMed]
- Ziv, Y., Burns, L. D., Cocker, E. D. et al.: Long-term dynamics of CA1 hippocampal place codes. Nat. Neurosci., 16, 264-266 (2013)[PubMed]
- Moita, M. A., Rosis, S., Zhou, Y. et al.: Putting fear in its place: remapping of hippocampal place cells during fear conditioning. J. Neurosci., 24, 7015-7023 (2004)[PubMed]
- Dupret, D., O'Neill, J., Pleydell-Bouverie, B. et al.: The reorganization and reactivation of hippocampal maps predict spatial memory performance. Nat. Neurosci., 13, 995-1002 (2010)[PubMed]
- Kaifosh, P., Lovett-Barron, M., Turi, G. F. et al.: Septo-hippocampal GABAergic signaling across multiple modalities in awake mice. Nat. Neurosci., 16, 1182-1184 (2013)[PubMed]
- Lovett-Barron, M., Kaifosh, P., Kheirbek, M. A. et al.: Dendritic inhibition in the hippocampus supports fear learning. Science, 343, 857-863 (2014)[PubMed]
- Wood, E. R., Dudchenko, P. A. & Eichenbaum, H.: The global record of memory in hippocampal neuronal activity. Nature, 397, 613-616 (1999)[PubMed]
- MacDonald, C. J., Lepage, K. Q., Eden, U. T. et al.: Hippocampal "time cells" bridge the gap in memory for discontiguous events. Neuron, 71, 737-749 (2011)[PubMed]
- Fyhn, M., Molden, S., Witter, M. P. et al.: Spatial representation in the entorhinal cortex. Science, 305, 1258-1264 (2004)[PubMed]
- Hafting, T., Fyhn, M., Molden, S. et al.: Microstructure of a spatial map in the entorhinal cortex. Nature, 436, 801-806 (2005)[PubMed]
- Boccara, C. N., Sargolini, F., Thoresen, V. H. et al.: Grid cells in pre- and parasubiculum. Nat. Neurosci., 13, 987-994 (2010)[PubMed]
- Barry, C., Hayman, R., Burgess, N. et al.: Experience-dependent rescaling of entorhinal grids. Nat. Neurosci., 10, 682-684 (2007)[PubMed]
- Barry, C., Ginzberg, L. L., O'Keefe, J. et al.: Grid cell firing patterns signal environmental novelty by expansion. Proc. Natl. Acad. Sci. USA, 109, 17687-17692 (2012)[PubMed]
- Taube, J. S., Muller, R. U. & Ranck, J. B. Jr.: Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. J. Neurosci., 10, 420-435 (1990)[PubMed]
- Taube, J. S., Muller, R. U. & Ranck, J. B. Jr.: Head-direction cells recorded from the postsubiculum in freely moving rats. II. Effects of environmental manipulations. J. Neurosci., 10, 436-447 (1990)[PubMed]
- Clark, B. J. & Taube, J. S.: Vestibular and attractor network basis of the head direction cell signal in subcortical circuits. Front. Neural Circuits, 6, 7 (2012)[PubMed]
- Lever, C., Burton, S., Jeewajee, A. et al.: Boundary vector cells in the subiculum of the hippocampal formation. J. Neurosci., 29, 9771-9777 (2009)[PubMed]
- Sargolini, F., Fyhn, M., Hafting, T. et al.: Conjunctive representation of position, direction, and velocity in entorhinal cortex. Science, 312, 758-762 (2006)[PubMed]
- Solstad, T., Boccara, C. N., Kropff, E. et al.: Representation of geometric borders in the entorhinal cortex. Science, 322, 1865-1868 (2008)[PubMed]
- Deshmukh, S. S. & Knierim, J. J.: Representation of non-spatial and spatial information in the lateral entorhinal cortex. Front. Behav. Neurosci., 5, 69 (2011)[PubMed]
- McNaughton, B. L., Battaglia, F. P., Jensen, O. et al.: Path integration and the neural basis of the 'cognitive map'. Nat. Rev. Neurosci., 7, 663-678 (2006)[PubMed]
- Solstad, T., Moser, E. I. & Einevoll, G. T.: From grid cells to place cells: a mathematical model. Hippocampus, 16, 1026-1031 (2006)[PubMed]
- Zhang, S. J., Ye, J., Miao, C. et al.: Optogenetic dissection of entorhinal-hippocampal functional connectivity. Science, 340, 1232627 (2013)[PubMed]
- Koenig, J., Linder, A. N., Leutgeb, J. K. et al.: The spatial periodicity of grid cells is not sustained during reduced theta oscillations. Science, 332, 592-595 (2011)[PubMed]
- Brandon, M. P., Koenig, J., Leutgeb, J. K. et al.: New and distinct hippocampal place codes are generated in a new environment during septal inactivation. Neuron, 82, 789-796 (2014)[PubMed]
- Hales, J. B., Schlesiger, M. I., Leutgeb, J. K. et al.: Medial entorhinal cortex lesions only partially disrupt hippocampal place cells and hippocampus-dependent place memory. Cell Rep., 9, 893-901 (2014)[PubMed]
- Bonnevie, T., Dunn, B., Fyhn, M. et al.: Grid cells require excitatory drive from the hippocampus. Nat. Neurosci., 16, 309-317 (2013)[PubMed]
- Kentros, C., Hargreaves, E., Hawkins, R. D. et al.: Abolition of long-term stability of new hippocampal place cell maps by NMDA receptor blockade. Science, 280, 2121-2126 (1998)[PubMed]
- McHugh, T. J., Blum, K. I., Tsien, J. Z. et al.: Impaired hippocampal representation of space in CA1-specific NMDAR1 knockout mice. Cell, 87, 1339-1349 (1996)[PubMed]
- Rotenberg, A., Mayford, M., Hawkins, R. D. et al.: Mice expressing activated CaMKII lack low frequency LTP and do not form stable place cells in the CA1 region of the hippocampus. Cell, 87, 1351-1361 (1996)[PubMed]
- Cho, Y. H., Giese, K. P., Tanila, H. et al.: Abnormal hippocampal spatial representations in αCaMKIIT286A and CREBαΔ- mice. Science, 279, 867-869 (1998)[PubMed]
- Rotenberg, A., Abel, T., Hawkins, R. D. et al.: Parallel instabilities of long-term potentiation, place cells, and learning caused by decreased protein kinase A activity. J. Neurosci., 20, 8096-8102 (2000)[PubMed]
- Resnik, E., McFarland, J. M., Sprengel, R. et al.: The effects of GluA1 deletion on the hippocampal population code for position. J. Neurosci., 32, 8952-8968 (2012)[PubMed]
- Lee, D., Lin, B. J. & Lee, A. K.: Hippocampal place fields emerge upon single-cell manipulation of excitability during behavior. Science, 337, 849-853 (2012)[PubMed]
- Sheffield, M. E. & Dombeck, D. A.: Calcium transient prevalence across the dendritic arbour predicts place field properties. Nature, 517, 200-204 (2015)[PubMed]
- Colgin, L. L., Denninger, T., Fyhn, M. et al.: Frequency of gamma oscillations routes flow of information in the hippocampus. Nature, 462, 353-357 (2009)[PubMed]
- Montgomery, S. M. & Buzsaki, G.: Gamma oscillations dynamically couple hippocampal CA3 and CA1 regions during memory task performance. Proc. Natl. Acad. Sci. USA, 104, 14495-14500 (2007)[PubMed]
- Yamamoto, J., Suh, J., Takeuchi, D. et al.: Successful execution of working memory linked to synchronized high-frequency gamma oscillations. Cell, 157, 845-857 (2014)[PubMed]
- Kemere, C., Carr, M. F., Karlsson, M. P. et al.: Rapid and continuous modulation of hippocampal network state during exploration of new places. PLoS One, 8, e73114 (2013)[PubMed]
- Chen, Z., Resnik, E., McFarland, J. M. et al.: Speed controls the amplitude and timing of the hippocampal gamma rhythm. PLoS One 6, e21408 (2011)[PubMed]
- Reijmers, L. G., Perkins, B. L., Matsuo, N. et al.: Localization of a stable neural correlate of associative memory. Science, 317, 1230-1233 (2007)[PubMed]
- Matsuo, N., Reijmers, L. & Mayford, M.: Spine-type-specific recruitment of newly synthesized AMPA receptors with learning. Science, 319, 1104-1107 (2008)[PubMed]
著者プロフィール
略歴:2009年 東京大学大学院薬学系研究科 修了,同年 ノルウェーNorwegian University of Science and Technology博士研究員,2013年 京都大学生命科学系キャリアパス形成ユニット 研究員を経て,2014年より大阪大学大学院医学系研究科 研究員.
研究テーマ:海馬および嗅内皮質における情報処理の機構.脳に流れる情報を手に取るように見たい.
松尾 直毅(Naoki Matsuo)
大阪大学大学院医学系研究科 独立准教授.
© 2015 北西卓磨・松尾直毅 Licensed under CC 表示 2.1 日本
