眼を動かしても世界が動かないのはなぜか
2015/09/10
北澤 茂
(大阪大学大学院生命機能研究科 ダイナミックブレインネットワーク研究室)
email:北澤 茂
領域融合レビュー, 4, e012 (2015) DOI: 10.7875/leading.author.4.e012
Shigeru Kitazawa: Why does the world remain stable while we move our eyes?

われわれはサッケードとよばれる1秒に3回もの高速な眼球の運動により外界の情報を得ている.しかし,網膜像のぶれに気づくことはなく,外界はつねに安定している.目を動かしても世界が動かないのはなぜだろうか.この問題は,少なくとも1000年前のアラビアの学術書にまでさかのぼることができ,以後,知の巨人たちが論考を重ねてきた.それらの論考の集大成は,サッケードの運動指令の遠心性コピーを使ってサッケードにより生じる網膜からの感覚のフィードバックが抑制される,というサッケード抑制の図式である.しかし,このモデルは問題のひとつ,なぜ網膜像のぶれが知覚されないのかを説明するにすぎない.網膜像がぶれているあいだにも視覚の途切れることはないことから,情報の空白をうめて意識される世界の連続性を保つしくみがあるはずである.つまり,2つ目の問題は,情報の空白が見えないのはなぜかと定義することができる.これらの問題がかたづいたとしても,目が動いた前後の網膜像がずれているという事実を変えることはできない.2枚の網膜像が異なるにもかかわらず,われわれの世界が微動だにしないのはなぜだろうか.ずれた網膜像はいかにして心の中の世界において同じ部分に対応づけられるのか,これが第3にして最大の問題である.このレビューでは,これら3つの問題についてこれまでに得られた知見をふりかえり,現時点でもっとも妥当と考えられる解について検討する.
われわれはサッケード(saccade)とよばれる1秒に3回もの高速な眼球の運動により外界の情報を得ている.しかし,網膜像のぶれには気づくことはなく,外界はつねに安定している.目を動かしても世界が動かないのはなぜだろうか.この問題は,少なくとも1000年前のアラビアの学術書にまでさかのぼることができ,以後,古今東西の科学と哲学につうじた知の巨人たちが論考を重ねてきた1-4).たとえば,目尻を軽く押すと世界がゆれることをはじめて書き残したのはデカルトである1).通常の眼球の運動ではまったくゆれないのに,指で押すと世界がゆれる.ということは,外眼筋を動かすための指令が視覚の世界の安定に寄与しているに違いない.運動指令の遠心性コピーという言葉とともに,現代の文献においてよく引用されるのは,サッケードの運動指令の遠心性コピーを使ってサッケードにより生じる網膜からの感覚のフィードバックが抑制される,というおよそ60年前にとなえられた図式である5)(図1).

突然,外眼筋が動かなくなった場合,この図式からは何が予測されるだろうか.感覚のフィードバックがなくなってしまうので,遠心性コピーの信号だけが脳により解釈され,世界は眼を動かそうとする意思の方向に動いて見えるはずである.早くも19世紀なかばには,急性の外眼筋の麻痺の症例において世界が目を動かそうとする意思の方向に動くことが報告されている3).1976年には,筋弛緩薬を用いて全身の筋肉をなかば弛緩させた状態において,覚醒したボランティアが“眼球の運動の意思”により世界が動くという経験を報告している6).この図式には根拠があるのだ.
しかし,遠心性コピーにもとづく網膜からの感覚のフィードバックの抑制だけでは解決できない問題もある.遠心性コピーにより動きの知覚を消すことできたとしても,サッケード中に視覚の情報が失われるという事実は消すことができない.情報が空白となる期間に,われわれの意識は何を見ているのだろうか.いい換えると,サッケードにより生じる情報の空白を脳はどのようにして補填しているのか.これが2番目の問題である.
さらに,目が動いている最中の2つの問題がかたづいたとしても,目が動いた前後の網膜像はずれているという事実が残る.2枚の網膜像が異なるにもかかわらず,われわれの世界が微動だにしないのはなぜだろうか.2枚の異なる網膜像は,それぞれ心の中の動かない世界の同じ部分に対応づけられているはずなのだ.ずれた網膜像はいかにして世界の同じ部分に対応づけられ統合されるのか,これが第3にして最大の問題である.
このレビューでは,これら3つの問題(図2)を,歴史的な経緯をふまえつつ,最新のデータと仮説にもとづいて検討する.

遠心性コピーにもとづくサッケード抑制を工学的に描くと,サッケードにより生じる網膜像のぶれの信号を予測して打ち消す,という図式が得られる(図1).この図式は,いささか乱暴ではあるが,サッケード中に網膜から入力する信号すべてを遮断してしまえば実現できる.本当にそのような完全な遮断が実現されているのだろうか.また,信号が遮断されているとすれば,脳のどこで遮断されているのだろうか.実際に,サッケード中に提示された光を感じる閾値は,眼が静止しているときに比べ上昇する(これが,狭義の“サッケード抑制”である).しかし,この抑制は限定的であり閾値は2~3倍になるすぎない7).2004年には,ヒトの網膜および1次視覚野を磁気による誘導電流により刺激し,眼の内部において閃光が生じる刺激の閾値が,サッケード中とサッケードのあとでどのように変化するかが調べられた8).その結果,サッケード中の網膜への刺激の閾値は2倍に上昇する一方,1次視覚野への刺激の閾値はまったく不変であることが明らかにされた.これらの結果は,網膜からの入力は網膜と1次視覚野とのあいだ(おそらく,外側膝状体のレベル)で若干の抑制をうけるものの,抑制は不完全で網膜からの信号を遮断するほどには強くないことを意味する.日常の光にあふれた環境においては,網膜からの信号はサッケード中にも1次視覚野に到達しているのだ.
しかし,1次視覚野のレベルでの活動が完全には遮断されていないとしても,動きを表現する大脳皮質の領域は強力なサッケード抑制をうけているかもしれない.動きに応じるニューロンは網膜,外側膝状体,1次視覚野に存在するが,サルにおいて動きを専門に表現する領域は上側頭溝の後方の中側頭野および内側上側頭野である.ヒトにも相同な領域が存在し,この領域に障害をうけた患者には,車が突然ワープして近づいてきたり,気がつくとコップの水があふれていたりといった不都合が生じる9).しかし,実際にこの領域の活動を調べてみると,サッケード中の活動の減少は限定的で十分な活動が維持されていた10).つまり,動きを表現する領域の活動もごく軽い抑制をうけているにすぎない.
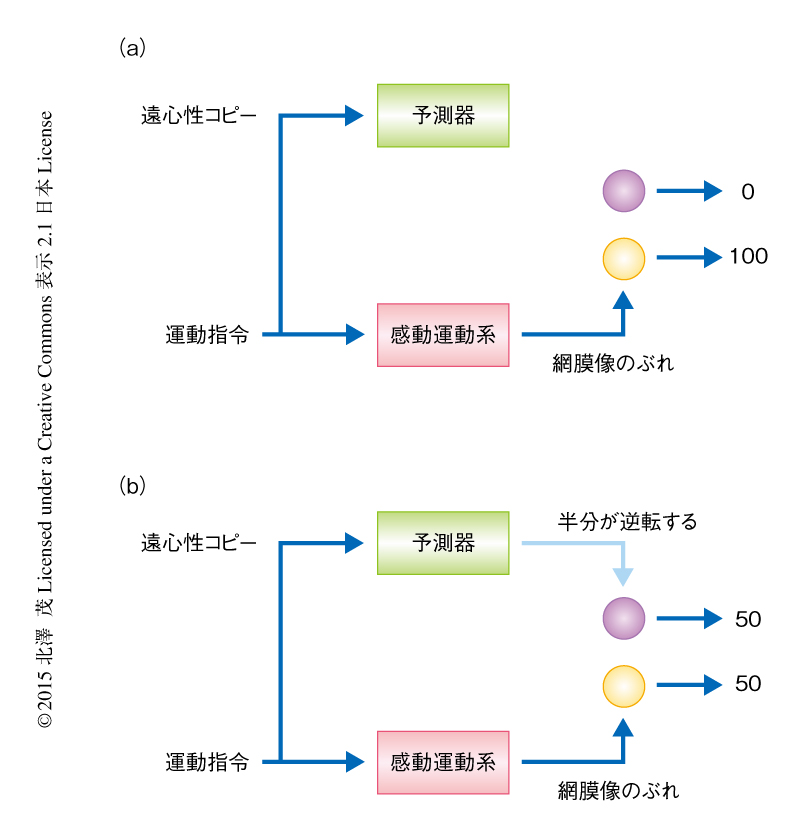
発火の頻度が高いままで,どのようにして動きの知覚を遮断するのだろうか.2002年,驚くべきニューロンの存在が報告された10).中側頭野および内側上側頭野のニューロンは受容野において特定の方向および速さをもつ動きの刺激に対しよく応答する.つまり,方向選択性をもつ.ところが,中側頭野および内側上側頭野の30~40%のニューロンの方向選択性がサッケード中に逆転したというのだ.じつは,ある方向の動きの知覚は,その方向に選択性をもつニューロンとその反対の方向に選択性をもつニューロンの活動の差分として生じると考えられている.この考えにもとづけば,半数のニューロンの方向選択性が逆転すれば,どのような向きにサッケードしようとも動きの知覚は中和されることになる.たとえば,左向きの動きに応じる左向きニューロンと右向きの動きに応じる右向きニューロンが100個ずつあるところ,右向きのサッケードが生じたと仮定しよう.もし,いかなる制御機構も存在しなければ,100個の左向きニューロンが活動する一方,右向きニューロンは活動しないので,左向きの知覚が生じる.つまり,世界が左向きに動いて見えるはずである(図3a).しかし,サッケードの際に半数のニューロンの方向選択性が逆転するとすれば,右向きのサッケードの際に活動する左向きニューロンの数は50個に半減する一方,50個の右向きニューロンは一時的に左向きを好むようになり活動する.その結果,左向きニューロンと右向きニューロンは同数になり動きの知覚は生じない(図3b).
筆者は,この中和説が実際のデータともっともよくあう説明であると感じている.しかし,脳はどうやってサッケードのたびにニューロンの方向選択性を逆転させてはもとにもどすなどという離れ業を演じているのだろうか.その謎は未解明の問題として残されている.
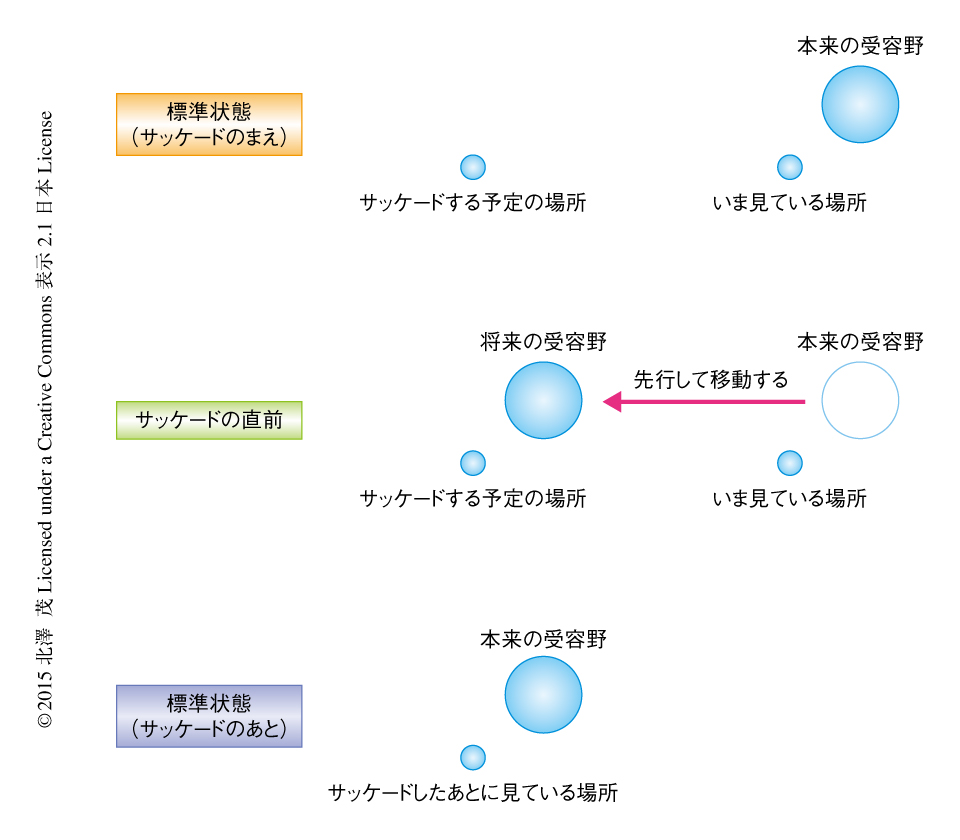
網膜像がぶれることにより生じる動きの知覚がいかにたくみに遮断あるいは中和されたとしても,サッケード中の網膜からの情報はぼけている.にもかかわらず,われわれの視覚の世界はいささかも曇ることはない.情報が途絶しているあいだ,いったい全体,われわれは何を見ているのだろうか.筆者がこの答えになると考えているのは,1992年に発見された受容野のリマッピングである11)(図4).頭頂葉におけるサッケードの中枢として知られている外側頭頂間野において,視覚応答性ニューロンの受容野が固視点の右上にあるとしよう.いま,サルがまさに左にサッケードしようとする瞬間に,このニューロンの受容野が,もとの受容野の位置からこれから生じるサッケードのベクトルの分だけ平行移動した位置に,一時的にリマッピングするというのだ.この場所はサッケードののちの本来の受容野の位置であるから,サッケードの直前には将来の受容野の位置をしめる.まとめると,サッケードの直前に外側頭頂間野のかなりのニューロンの受容野が,将来の受容野へと飛ぶのだ.つまり,外側頭頂間野のニューロンは,サッケードの直前からサッケードのあとの活動の状態を予測していることになる.サッケード中の情報の欠損は予測的に補填されているらしい.
このような受容野のリマッピングは,前頭葉におけるサッケードの中枢である前頭眼野でも生じている.しかも,このリマッピングは上丘から視床を経由して前頭眼野にいたる経路を視床において遮断すると生じなくなる.つまり,上丘からの遠心性コピーが視床を経由して入力することによりリマッピングが生じるというところまでわかっている12).しかし,どのようにしてこのようなリマッピングが可能になるのか,という肝心の機構は推測の域をでない.
このリマッピングこそが視覚の世界を安定化させるための主要な機構であり,のちに述べる外部の座標系などは不要であると考えている研究者もいるが13),その主張の根拠は疑問視されている14).筆者は,リマッピングをサッケード中に失われる情報の補填の機構として位置づけるのが妥当だと考えている.
3番目にして最大の問題について考えよう.サッケード中の動きの情報を中和し,サッケードの直前に取得した将来の受容野の情報により補填したとしても,サッケードの前後でサッケードのベクトルの分だけずれた2枚の網膜像が残ることに変わりはない.これら2枚の網膜像は,われわれの意識のなかに存在している(と思われる)不動の外界の同じ場所に写像されなければならない.
網膜の2次元のトポロジーは,ほぼそのまま,1次視覚野,2次視覚野をへて,3次視覚野,中側頭野などのいわゆる視覚の背側系や,4次視覚野などの腹側系へとひきつがれていく.これらの領域のニューロンは網膜の特定の領域を刺激すると活動する.つまり,これらのニューロンは網膜の座標系をひきずっている.不動の外界が脳のなかに表現されているとすれば,その領域のニューロンはどのような活動を示すべきだろうか.目がどこをむいていても,そして,頭がどこをむいていても,空間のある場所に刺激をあたえると応答する,そんなニューロンには十分な資格があるだろう.
1985年,頭頂葉において,受容野の位置は網膜の座標系により固定されているものの,眼球の位置に応じて活動のゲインが制御をうけるゲインフィールドニューロンが発見された15).これらのニューロンの情報を組み合わせれば,頭部に固定した座標系のどこに刺激が位置するかを計算することができる.さらに1993年,頭頂葉の内側後方に位置する頭頂後頭野に,ゲインフィールドニューロンのほかに,目がどこをむいていてもスクリーンの同じ場所に受容野が固定されて動かないという頭蓋座標系ニューロンの存在することが報告された16).そののち,腹側頭頂間野にも,眼球の位置に応じて活動を変化させるゲインフィールドニューロンおよび頭蓋座標系ニューロンの存在することが報告された17).
この路線を延長すれば,体幹に対する頭部の位置に応じたゲインフィールドをもつニューロンがあれば,肩に固定した座標系も構築することができそうである.しかし,体の位置によらない本当の外部空間の座標系を構築するには,外界に対する体の位置に応じたゲインフィールドが必要になる.外界に対する自己の位置を計るには外部の座標系が必要だから,この論法は循環論法におちいってしまう.ゲインフィールドを拡張して構築することのできるのは,体に固定された自己中心の座標系に限られるように思われる.さらに,このような拡張にともなう問題は,ゲインフィールドの階層を重ねるごとに計算量および誤差が積みあがっていくことである.1秒に3回も急速に生じる眼球の位置の変化に応じて,誤差なく網膜像を上位の座標系にマッピングするのは至難の業だろう.
これらの困難を解決することのできる座標系として,近年,急速に関心をあつめているのが,自己を離れた外界の他者を基準とする他者中心の座標系である.従来から,意識してある程度の時間をかけて努力すれば目標の位置を他者中心の座標系により記憶することのできることは知られていた.しかし,1秒間に3回生じるサッケードの際に役だつには意識的な努力は不都合であり,他者中心の座標系が無意識かつ高速に動作する必要がある.最近,あいついで報告された研究によれば,背景座標系は瞬時かつ自動的に目標の位置を表現することができるようだ18-20).たとえば,四角の背景のある画面のなかで標的となる赤色の線分の傾きの方位を回答する課題において,標的の現われる位置に事前にキュー刺激をあたえることにより反応の速度は速くなるかどうか,また,正答率は上がるかどうかが調べられた18).その結果,四角の位置が網膜のうえでずれたとしても,四角に対して同じ位置(背景座標系において標的と同じ位置)にキュー刺激をあたえると,反応が速くなり正答率も高くなる効果のあることが示された.
最近の機能的磁気共鳴画像(fMRI)法を用いた研究により,背景座標系は右の楔前部に存在することが明らかにされた21).この領域は,サルにおいて頭部に固定された座標系の発見された頭頂後頭野の,ヒトにおける相同部位の直前に位置している.つまり,後ろ(1次視覚野)から前にむかって網膜座標系-頭部中心座標系-背景座標系という座標系の変換が進行していく可能性がある.また,楔前部は脳のネットワーク解析において注目されるハブ領域のひとつであり22),“世界”を統一するには都合のよい場所であることを付言する.
筆者らを含めた複数の研究グループは,視覚の安定性に決定的な貢献をしているのは背景(座標系)であると主張している14,19,21).間接的な状況証拠をいくつかあげる.1)筋弛緩薬を用いた非動化の実験では,暗闇で目を動かそうとすると光点が動いたのに対し,やや明るくして部屋の背景が見える状態にすると光点はまったく動かなくなった23).2)網膜のうえの残像は,1点だけであればサッケードにともなって動くように見えたが,複雑な背景の残像はまったく動かなかった24).3)サッケード中に13個の物体をまとめて移動すると,4度も動かしても半数の試行では動いたことに気がつかなかった25).これらの知見は,サッケードの前後の2枚の網膜像が背景を基準とした座標系に収斂することを示唆する.
ここで,おそらくつぎのような疑問がうかぶのではないか.デカルトが記載した,目を指でくり返し押す操作により背景が動いて見えるのはなぜだろう.ずれた網膜像が背景座標系に収斂するなら,眼を指で押しても世界は動かないはずではないか.じつは,1958年に興味深い現象が報告されている26).オシロスコープの持続的な輝点は目尻を指でくり返し押すと連続して動いたが,同時にストロボ光を5~6 Hzの周波数で光らせると,ストロボ光により断続的に照明されたオシロスコープの枠を含めた部屋の背景はまったく動かず,そのなかでオシロスコープの輝点だけが動いて見えたという.つまり,断続的な背景の情報は網膜のうえで位置が変わっても不動の背景としてまとめられる一方,網膜のうえで連続的に移動する光跡はそのまま動きとして知覚されることになる.
背景座標系の存在を結論した最近の研究はすべて,30~300ミリ秒の短い空白をはさんで背景の映像を網膜のうえの異なる位置に投影している18-20).背景を基準にして2枚の網膜像を統一する機構はサッケードそのものにより駆動されるのではなく,動きの信号が短時間のあいだ遮断されることにより駆動されるのではないか.サッケードの前後では動きの情報が中和されて遮断されることにより,その前後の網膜像から背景が抽出され同一視される,と考えれば過去の知見のつじつまがあう.
冒頭に述べた3つの問いに対する,現在,得られているデータにもとづいた暫定の解は以下のとおりである.1)動きの情報はサッケード中に動きに応答するニューロンの方向選択性を制御することにより中和されている.2)サッケードにともなう情報の空白は受容野の予測的なリマッピングにより補填されている.3)2枚のずれた網膜像は情報の短時間の途絶により駆動される背景座標系に統合される.もちろん,問いのたて方もその暫定の解も定説ではないので,今後,大きな修正をうける可能性がある.しかし,1000年にわたり知の巨人たちが挑んできた問題に現代の神経科学が最終的な解答をあたえるのは,それほど遠い未来のことではないだろう.
略歴:1991年 東京大学大学院医学系研究科 修了,1993年 東京大学医学部 助手,1994年 工業技術院電子技術総合研究所 研究員,1995年 同 主任研究官,2001年 産業技術総合研究所脳神経情報研究部門 主任研究員,2003年 順天堂大学医学部 教授を経て,2011年より大阪大学大学院生命機能研究科 教授.
研究テーマ:運動制御の最適化の神経機構.空間と時間の知覚の神経科学.
関心事:目を動かしても世界が動かないのはなぜかという問いに納得できる答えをだすこと.他方,信号の時間順序の判断が入れ替わる錯覚に注目して“こころ”が感じる時間の神経基盤の解明にも取り組んでいる.空間知覚の安定性を優先するために時間の流れにしわよせがいく,という考えで研究を進めている.
研究室URL:http://www.fbs.osaka-u.ac.jp/jpn/general/lab/181/
© 2015 北澤 茂 Licensed under CC 表示 2.1 日本
(大阪大学大学院生命機能研究科 ダイナミックブレインネットワーク研究室)
email:北澤 茂
領域融合レビュー, 4, e012 (2015) DOI: 10.7875/leading.author.4.e012
Shigeru Kitazawa: Why does the world remain stable while we move our eyes?
要 約
われわれはサッケードとよばれる1秒に3回もの高速な眼球の運動により外界の情報を得ている.しかし,網膜像のぶれに気づくことはなく,外界はつねに安定している.目を動かしても世界が動かないのはなぜだろうか.この問題は,少なくとも1000年前のアラビアの学術書にまでさかのぼることができ,以後,知の巨人たちが論考を重ねてきた.それらの論考の集大成は,サッケードの運動指令の遠心性コピーを使ってサッケードにより生じる網膜からの感覚のフィードバックが抑制される,というサッケード抑制の図式である.しかし,このモデルは問題のひとつ,なぜ網膜像のぶれが知覚されないのかを説明するにすぎない.網膜像がぶれているあいだにも視覚の途切れることはないことから,情報の空白をうめて意識される世界の連続性を保つしくみがあるはずである.つまり,2つ目の問題は,情報の空白が見えないのはなぜかと定義することができる.これらの問題がかたづいたとしても,目が動いた前後の網膜像がずれているという事実を変えることはできない.2枚の網膜像が異なるにもかかわらず,われわれの世界が微動だにしないのはなぜだろうか.ずれた網膜像はいかにして心の中の世界において同じ部分に対応づけられるのか,これが第3にして最大の問題である.このレビューでは,これら3つの問題についてこれまでに得られた知見をふりかえり,現時点でもっとも妥当と考えられる解について検討する.
はじめに
われわれはサッケード(saccade)とよばれる1秒に3回もの高速な眼球の運動により外界の情報を得ている.しかし,網膜像のぶれには気づくことはなく,外界はつねに安定している.目を動かしても世界が動かないのはなぜだろうか.この問題は,少なくとも1000年前のアラビアの学術書にまでさかのぼることができ,以後,古今東西の科学と哲学につうじた知の巨人たちが論考を重ねてきた1-4).たとえば,目尻を軽く押すと世界がゆれることをはじめて書き残したのはデカルトである1).通常の眼球の運動ではまったくゆれないのに,指で押すと世界がゆれる.ということは,外眼筋を動かすための指令が視覚の世界の安定に寄与しているに違いない.運動指令の遠心性コピーという言葉とともに,現代の文献においてよく引用されるのは,サッケードの運動指令の遠心性コピーを使ってサッケードにより生じる網膜からの感覚のフィードバックが抑制される,というおよそ60年前にとなえられた図式である5)(図1).
突然,外眼筋が動かなくなった場合,この図式からは何が予測されるだろうか.感覚のフィードバックがなくなってしまうので,遠心性コピーの信号だけが脳により解釈され,世界は眼を動かそうとする意思の方向に動いて見えるはずである.早くも19世紀なかばには,急性の外眼筋の麻痺の症例において世界が目を動かそうとする意思の方向に動くことが報告されている3).1976年には,筋弛緩薬を用いて全身の筋肉をなかば弛緩させた状態において,覚醒したボランティアが“眼球の運動の意思”により世界が動くという経験を報告している6).この図式には根拠があるのだ.
しかし,遠心性コピーにもとづく網膜からの感覚のフィードバックの抑制だけでは解決できない問題もある.遠心性コピーにより動きの知覚を消すことできたとしても,サッケード中に視覚の情報が失われるという事実は消すことができない.情報が空白となる期間に,われわれの意識は何を見ているのだろうか.いい換えると,サッケードにより生じる情報の空白を脳はどのようにして補填しているのか.これが2番目の問題である.
さらに,目が動いている最中の2つの問題がかたづいたとしても,目が動いた前後の網膜像はずれているという事実が残る.2枚の網膜像が異なるにもかかわらず,われわれの世界が微動だにしないのはなぜだろうか.2枚の異なる網膜像は,それぞれ心の中の動かない世界の同じ部分に対応づけられているはずなのだ.ずれた網膜像はいかにして世界の同じ部分に対応づけられ統合されるのか,これが第3にして最大の問題である.
このレビューでは,これら3つの問題(図2)を,歴史的な経緯をふまえつつ,最新のデータと仮説にもとづいて検討する.
1.網膜像のぶれの知覚をどのように抑制しているのか
遠心性コピーにもとづくサッケード抑制を工学的に描くと,サッケードにより生じる網膜像のぶれの信号を予測して打ち消す,という図式が得られる(図1).この図式は,いささか乱暴ではあるが,サッケード中に網膜から入力する信号すべてを遮断してしまえば実現できる.本当にそのような完全な遮断が実現されているのだろうか.また,信号が遮断されているとすれば,脳のどこで遮断されているのだろうか.実際に,サッケード中に提示された光を感じる閾値は,眼が静止しているときに比べ上昇する(これが,狭義の“サッケード抑制”である).しかし,この抑制は限定的であり閾値は2~3倍になるすぎない7).2004年には,ヒトの網膜および1次視覚野を磁気による誘導電流により刺激し,眼の内部において閃光が生じる刺激の閾値が,サッケード中とサッケードのあとでどのように変化するかが調べられた8).その結果,サッケード中の網膜への刺激の閾値は2倍に上昇する一方,1次視覚野への刺激の閾値はまったく不変であることが明らかにされた.これらの結果は,網膜からの入力は網膜と1次視覚野とのあいだ(おそらく,外側膝状体のレベル)で若干の抑制をうけるものの,抑制は不完全で網膜からの信号を遮断するほどには強くないことを意味する.日常の光にあふれた環境においては,網膜からの信号はサッケード中にも1次視覚野に到達しているのだ.
しかし,1次視覚野のレベルでの活動が完全には遮断されていないとしても,動きを表現する大脳皮質の領域は強力なサッケード抑制をうけているかもしれない.動きに応じるニューロンは網膜,外側膝状体,1次視覚野に存在するが,サルにおいて動きを専門に表現する領域は上側頭溝の後方の中側頭野および内側上側頭野である.ヒトにも相同な領域が存在し,この領域に障害をうけた患者には,車が突然ワープして近づいてきたり,気がつくとコップの水があふれていたりといった不都合が生じる9).しかし,実際にこの領域の活動を調べてみると,サッケード中の活動の減少は限定的で十分な活動が維持されていた10).つまり,動きを表現する領域の活動もごく軽い抑制をうけているにすぎない.
発火の頻度が高いままで,どのようにして動きの知覚を遮断するのだろうか.2002年,驚くべきニューロンの存在が報告された10).中側頭野および内側上側頭野のニューロンは受容野において特定の方向および速さをもつ動きの刺激に対しよく応答する.つまり,方向選択性をもつ.ところが,中側頭野および内側上側頭野の30~40%のニューロンの方向選択性がサッケード中に逆転したというのだ.じつは,ある方向の動きの知覚は,その方向に選択性をもつニューロンとその反対の方向に選択性をもつニューロンの活動の差分として生じると考えられている.この考えにもとづけば,半数のニューロンの方向選択性が逆転すれば,どのような向きにサッケードしようとも動きの知覚は中和されることになる.たとえば,左向きの動きに応じる左向きニューロンと右向きの動きに応じる右向きニューロンが100個ずつあるところ,右向きのサッケードが生じたと仮定しよう.もし,いかなる制御機構も存在しなければ,100個の左向きニューロンが活動する一方,右向きニューロンは活動しないので,左向きの知覚が生じる.つまり,世界が左向きに動いて見えるはずである(図3a).しかし,サッケードの際に半数のニューロンの方向選択性が逆転するとすれば,右向きのサッケードの際に活動する左向きニューロンの数は50個に半減する一方,50個の右向きニューロンは一時的に左向きを好むようになり活動する.その結果,左向きニューロンと右向きニューロンは同数になり動きの知覚は生じない(図3b).
筆者は,この中和説が実際のデータともっともよくあう説明であると感じている.しかし,脳はどうやってサッケードのたびにニューロンの方向選択性を逆転させてはもとにもどすなどという離れ業を演じているのだろうか.その謎は未解明の問題として残されている.
2.情報の空白をいかに補填しているのか
網膜像がぶれることにより生じる動きの知覚がいかにたくみに遮断あるいは中和されたとしても,サッケード中の網膜からの情報はぼけている.にもかかわらず,われわれの視覚の世界はいささかも曇ることはない.情報が途絶しているあいだ,いったい全体,われわれは何を見ているのだろうか.筆者がこの答えになると考えているのは,1992年に発見された受容野のリマッピングである11)(図4).頭頂葉におけるサッケードの中枢として知られている外側頭頂間野において,視覚応答性ニューロンの受容野が固視点の右上にあるとしよう.いま,サルがまさに左にサッケードしようとする瞬間に,このニューロンの受容野が,もとの受容野の位置からこれから生じるサッケードのベクトルの分だけ平行移動した位置に,一時的にリマッピングするというのだ.この場所はサッケードののちの本来の受容野の位置であるから,サッケードの直前には将来の受容野の位置をしめる.まとめると,サッケードの直前に外側頭頂間野のかなりのニューロンの受容野が,将来の受容野へと飛ぶのだ.つまり,外側頭頂間野のニューロンは,サッケードの直前からサッケードのあとの活動の状態を予測していることになる.サッケード中の情報の欠損は予測的に補填されているらしい.
このような受容野のリマッピングは,前頭葉におけるサッケードの中枢である前頭眼野でも生じている.しかも,このリマッピングは上丘から視床を経由して前頭眼野にいたる経路を視床において遮断すると生じなくなる.つまり,上丘からの遠心性コピーが視床を経由して入力することによりリマッピングが生じるというところまでわかっている12).しかし,どのようにしてこのようなリマッピングが可能になるのか,という肝心の機構は推測の域をでない.
このリマッピングこそが視覚の世界を安定化させるための主要な機構であり,のちに述べる外部の座標系などは不要であると考えている研究者もいるが13),その主張の根拠は疑問視されている14).筆者は,リマッピングをサッケード中に失われる情報の補填の機構として位置づけるのが妥当だと考えている.
3.ずれた2枚の網膜像を統合する機構
3番目にして最大の問題について考えよう.サッケード中の動きの情報を中和し,サッケードの直前に取得した将来の受容野の情報により補填したとしても,サッケードの前後でサッケードのベクトルの分だけずれた2枚の網膜像が残ることに変わりはない.これら2枚の網膜像は,われわれの意識のなかに存在している(と思われる)不動の外界の同じ場所に写像されなければならない.
網膜の2次元のトポロジーは,ほぼそのまま,1次視覚野,2次視覚野をへて,3次視覚野,中側頭野などのいわゆる視覚の背側系や,4次視覚野などの腹側系へとひきつがれていく.これらの領域のニューロンは網膜の特定の領域を刺激すると活動する.つまり,これらのニューロンは網膜の座標系をひきずっている.不動の外界が脳のなかに表現されているとすれば,その領域のニューロンはどのような活動を示すべきだろうか.目がどこをむいていても,そして,頭がどこをむいていても,空間のある場所に刺激をあたえると応答する,そんなニューロンには十分な資格があるだろう.
1985年,頭頂葉において,受容野の位置は網膜の座標系により固定されているものの,眼球の位置に応じて活動のゲインが制御をうけるゲインフィールドニューロンが発見された15).これらのニューロンの情報を組み合わせれば,頭部に固定した座標系のどこに刺激が位置するかを計算することができる.さらに1993年,頭頂葉の内側後方に位置する頭頂後頭野に,ゲインフィールドニューロンのほかに,目がどこをむいていてもスクリーンの同じ場所に受容野が固定されて動かないという頭蓋座標系ニューロンの存在することが報告された16).そののち,腹側頭頂間野にも,眼球の位置に応じて活動を変化させるゲインフィールドニューロンおよび頭蓋座標系ニューロンの存在することが報告された17).
この路線を延長すれば,体幹に対する頭部の位置に応じたゲインフィールドをもつニューロンがあれば,肩に固定した座標系も構築することができそうである.しかし,体の位置によらない本当の外部空間の座標系を構築するには,外界に対する体の位置に応じたゲインフィールドが必要になる.外界に対する自己の位置を計るには外部の座標系が必要だから,この論法は循環論法におちいってしまう.ゲインフィールドを拡張して構築することのできるのは,体に固定された自己中心の座標系に限られるように思われる.さらに,このような拡張にともなう問題は,ゲインフィールドの階層を重ねるごとに計算量および誤差が積みあがっていくことである.1秒に3回も急速に生じる眼球の位置の変化に応じて,誤差なく網膜像を上位の座標系にマッピングするのは至難の業だろう.
これらの困難を解決することのできる座標系として,近年,急速に関心をあつめているのが,自己を離れた外界の他者を基準とする他者中心の座標系である.従来から,意識してある程度の時間をかけて努力すれば目標の位置を他者中心の座標系により記憶することのできることは知られていた.しかし,1秒間に3回生じるサッケードの際に役だつには意識的な努力は不都合であり,他者中心の座標系が無意識かつ高速に動作する必要がある.最近,あいついで報告された研究によれば,背景座標系は瞬時かつ自動的に目標の位置を表現することができるようだ18-20).たとえば,四角の背景のある画面のなかで標的となる赤色の線分の傾きの方位を回答する課題において,標的の現われる位置に事前にキュー刺激をあたえることにより反応の速度は速くなるかどうか,また,正答率は上がるかどうかが調べられた18).その結果,四角の位置が網膜のうえでずれたとしても,四角に対して同じ位置(背景座標系において標的と同じ位置)にキュー刺激をあたえると,反応が速くなり正答率も高くなる効果のあることが示された.
最近の機能的磁気共鳴画像(fMRI)法を用いた研究により,背景座標系は右の楔前部に存在することが明らかにされた21).この領域は,サルにおいて頭部に固定された座標系の発見された頭頂後頭野の,ヒトにおける相同部位の直前に位置している.つまり,後ろ(1次視覚野)から前にむかって網膜座標系-頭部中心座標系-背景座標系という座標系の変換が進行していく可能性がある.また,楔前部は脳のネットワーク解析において注目されるハブ領域のひとつであり22),“世界”を統一するには都合のよい場所であることを付言する.
筆者らを含めた複数の研究グループは,視覚の安定性に決定的な貢献をしているのは背景(座標系)であると主張している14,19,21).間接的な状況証拠をいくつかあげる.1)筋弛緩薬を用いた非動化の実験では,暗闇で目を動かそうとすると光点が動いたのに対し,やや明るくして部屋の背景が見える状態にすると光点はまったく動かなくなった23).2)網膜のうえの残像は,1点だけであればサッケードにともなって動くように見えたが,複雑な背景の残像はまったく動かなかった24).3)サッケード中に13個の物体をまとめて移動すると,4度も動かしても半数の試行では動いたことに気がつかなかった25).これらの知見は,サッケードの前後の2枚の網膜像が背景を基準とした座標系に収斂することを示唆する.
ここで,おそらくつぎのような疑問がうかぶのではないか.デカルトが記載した,目を指でくり返し押す操作により背景が動いて見えるのはなぜだろう.ずれた網膜像が背景座標系に収斂するなら,眼を指で押しても世界は動かないはずではないか.じつは,1958年に興味深い現象が報告されている26).オシロスコープの持続的な輝点は目尻を指でくり返し押すと連続して動いたが,同時にストロボ光を5~6 Hzの周波数で光らせると,ストロボ光により断続的に照明されたオシロスコープの枠を含めた部屋の背景はまったく動かず,そのなかでオシロスコープの輝点だけが動いて見えたという.つまり,断続的な背景の情報は網膜のうえで位置が変わっても不動の背景としてまとめられる一方,網膜のうえで連続的に移動する光跡はそのまま動きとして知覚されることになる.
背景座標系の存在を結論した最近の研究はすべて,30~300ミリ秒の短い空白をはさんで背景の映像を網膜のうえの異なる位置に投影している18-20).背景を基準にして2枚の網膜像を統一する機構はサッケードそのものにより駆動されるのではなく,動きの信号が短時間のあいだ遮断されることにより駆動されるのではないか.サッケードの前後では動きの情報が中和されて遮断されることにより,その前後の網膜像から背景が抽出され同一視される,と考えれば過去の知見のつじつまがあう.
おわりに
冒頭に述べた3つの問いに対する,現在,得られているデータにもとづいた暫定の解は以下のとおりである.1)動きの情報はサッケード中に動きに応答するニューロンの方向選択性を制御することにより中和されている.2)サッケードにともなう情報の空白は受容野の予測的なリマッピングにより補填されている.3)2枚のずれた網膜像は情報の短時間の途絶により駆動される背景座標系に統合される.もちろん,問いのたて方もその暫定の解も定説ではないので,今後,大きな修正をうける可能性がある.しかし,1000年にわたり知の巨人たちが挑んできた問題に現代の神経科学が最終的な解答をあたえるのは,それほど遠い未来のことではないだろう.
文 献
- Medendorp, W. P.: Spatial constancy mechanisms in motor control. Philos. Trans. R. Soc. Lond. B Biol. Sci., 366, 476-491 (2011)[PubMed]
- MacKay, D. M.: Visual stability and voluntary eye movement. in Central Processing of Visual Information A (Jung, R. ed.), pp.307-331, Springer-Verlag, Berlin (1973)
- Bridgeman, B., Van der Heijden, A. H. C. & Velichkovsky, B. M.: A theory of visual stability across saccadic eye movements. Behav. Brain Sci., 17, 247-258 (1994)
- Melcher, D.: Visual stability. Philos. Trans. R. Soc. Lond. B Biol. Sci., 366, 468-475 (2011)[PubMed]
- von Holst, E. & Mittelstaedt, H.: Das reafferenz princip: wedlselwirkungen zwishchen Zentrainervensystem und Peripherie. Die Naturwissenschaften, 37, 464-476 (1950)
- Stevens, J. K., Emerson, R. C., Gerstein, G. L. et al.: Paralysis of the awake human: visual perceptions. Vision Res., 16, 93-98 (1976)[PubMed]
- Campbell, F. W. & Wurtz, R. H.: Saccadic omission: why we do not see a grey-out during a saccadic eye movement. Vision Res., 18, 1297-1303 (1978)[PubMed]
- Thilo, K. V., Santoro, L., Walsh, V. et al.: The site of saccadic suppression. Nat. Neurosci., 7, 13-14 (2004)[PubMed]
- Zihl, J., von Cramon, D. & Mai, N.: Selective disturbance of movement vision after bilateral brain damage. Brain, 106(Pt.2), 313-340 (1983)[PubMed]
- Thiele, A., Henning, P., Kubischik, M. et al.: Neural mechanisms of saccadic suppression. Science, 295, 2460-2462 (2002)[PubMed]
- Duhamel, J. R., Colby, C. L. & Goldberg, M. E.: The updating of the representation of visual space in parietal cortex by intended eye movements. Science, 255, 90-92 (1992)[PubMed]
- Sommer, M. A. & Wurtz, R. H.: Visual perception and corollary discharge. Perception, 37, 408-418 (2008)[PubMed]
- Wurtz, R. H.: Neuronal mechanisms of visual stability. Vision Res., 48, 2070-2089 (2008)[PubMed]
- Bays, P. M. & Husain, M.: Spatial remapping of the visual world across saccades. Neuroreport, 18, 1207-1213 (2007)[PubMed]
- Andersen, R. A., Essick, G. K. & Siegel, R. M.: Encoding of spatial location by posterior parietal neurons. Science, 230, 456-458 (1985)[PubMed]
- Galletti, C., Battaglini, P. P. & Fattori, P.: Parietal neurons encoding spatial locations in craniotopic coordinates. Exp. Brain Res., 96, 221-229 (1993)[PubMed]
- Duhamel, J. R., Bremmer, F., Ben Hamed, S. et al.: Spatial invariance of visual receptive fields in parietal cortex neurons. Nature, 389, 845-848 (1997)[PubMed]
- Boi, M., Vergeer, M., Ogmen, H. et al.: Nonretinotopic exogenous attention. Curr. Biol., 21, 1732-1737 (2011)[PubMed]
- Lin, Z. & He, S.: Automatic frame-centered object representation and integration revealed by iconic memory, visual priming, and backward masking. J. Vis., 12, 24 (2012)[PubMed]
- Uchimura, M. & Kitazawa, S.: Cancelling prism adaptation by a shift of background: a novel utility of allocentric coordinates for extracting motor errors. J. Neurosci., 33, 7595-7602 (2013)[PubMed]
- Uchimura, M., Nakano, T., Morito, Y. et al.: Automatic representation of a visual stimulus relative to a background in the right precuneus. Eur. J. Neurosci., 42, 1651-1659 (2015)[PubMed]
- Hagmann, P., Cammoun, L., Gigandet, X. et al.: Mapping the structural core of human cerebral cortex. PLoS Biol., 6, e159 (2008)[PubMed]
- Matin, L., Picoult, E., Stevens, J. K. et al.: Oculoparalytic illusion: visual-field dependent spatial mislocalizations by humans partially paralyzed with curare. Science, 216, 198-201 (1982)[PubMed]
- Pelz, J. B. & Hayhoe, M. M.: The role of exocentric reference frames in the perception of visual direction. Vision Res., 35, 2267-2275 (1995)[PubMed]
- Bridgeman, B., Hendry, D. & Stark, L.: Failure to detect displacement of the visual world during saccadic eye movements. Vision Res., 15, 719-722 (1975)[PubMed]
- Mackay, D. M.: Perceptual stability of a stroboscopically lit visual field containing self-luminous objects. Nature, 181, 507-508 (1958)[PubMed]
著者プロフィール
略歴:1991年 東京大学大学院医学系研究科 修了,1993年 東京大学医学部 助手,1994年 工業技術院電子技術総合研究所 研究員,1995年 同 主任研究官,2001年 産業技術総合研究所脳神経情報研究部門 主任研究員,2003年 順天堂大学医学部 教授を経て,2011年より大阪大学大学院生命機能研究科 教授.
研究テーマ:運動制御の最適化の神経機構.空間と時間の知覚の神経科学.
関心事:目を動かしても世界が動かないのはなぜかという問いに納得できる答えをだすこと.他方,信号の時間順序の判断が入れ替わる錯覚に注目して“こころ”が感じる時間の神経基盤の解明にも取り組んでいる.空間知覚の安定性を優先するために時間の流れにしわよせがいく,という考えで研究を進めている.
研究室URL:http://www.fbs.osaka-u.ac.jp/jpn/general/lab/181/
© 2015 北澤 茂 Licensed under CC 表示 2.1 日本
