Wntシグナルの研究を基盤とした新規の抗がん剤の開発
2018/12/27
松本真司・菊池 章
(大阪大学大学院医学系研究科 分子病態生化学)
email:松本真司,菊池 章
領域融合レビュー, 7, e009 (2018) DOI: 10.7875/leading.author.7.e009
Shinji Matsumoto & Akira Kikuchi: Wnt signaling pathways in cancers: key targets and implications in cancer therapy.

Wntは分泌性糖タンパク質であり,Wntシグナル伝達経路を活性化することにより,増殖,分化,細胞運動,極性など,多岐にわたる細胞の応答を制御する.Wntシグナルは動物の胎生期における発生の過程に必須であり,種々の臓器の形成を制御する.出生ののちには臓器の恒常性の維持に関与し,再生や修復の過程において活性化される.一方,大腸がんをはじめとするヒトの種々のがんにおいて,Wntシグナル伝達経路の異常な活性化の関与について報告されている.これまで,Wntシグナル伝達経路の遮断を目的とした抗がん剤の開発が試みられてきたものの,医薬品として実用化された例はない.しかし,近年のヒトのがんゲノムの網羅的な解析やオルガノイド培養技術とあいまって,新たながんシグナルタンパク質の発見および分子標的治療薬の開発が期待されている.
1982年,マウスの乳がんの原因遺伝子としてInt-1遺伝子がクローニングされ,ショウジョウバエにおけるInt-1遺伝子のホモログがWingless遺伝子という分節の形成に重要な役割をはたす遺伝子であったことから,Int-1はWnt-1(Wingless + Int-1)と名づけられた1).それから35年以上がたち,腫瘍医学,発生生物学,幹細胞生物学においても解析が進み,Wntの研究領域は大きく展開してきた.ヒトのがんにおけるWntシグナルの重要性は,おもに大腸がんにおいて明らかにされてきた.しかし,近年のヒトのがんゲノムの網羅的な解析や,幹細胞の研究において発展したオルガノイド培養技術の応用などにより,Wntシグナル伝達経路の構成タンパク質およびWnt関連タンパク質の異常がヒトの多様ながんに関与することが明らかにされてきた.一方で,これまで,Wntシグナル伝達経路の遮断を目的とした抗がん剤の開発が試みられてきたものの,効果と安全性の問題から医薬品として実用化された例はない.このレビューにおいては,これまでに明らかにされてきたWntシグナルの活性化の機構に関する知見から,ヒトのがんにおけるWntシグナル伝達経路の構成タンパク質の異常,また,現在,臨床試験が進められているWnt関連タンパク質を標的にした抗がん剤の開発について解説する.
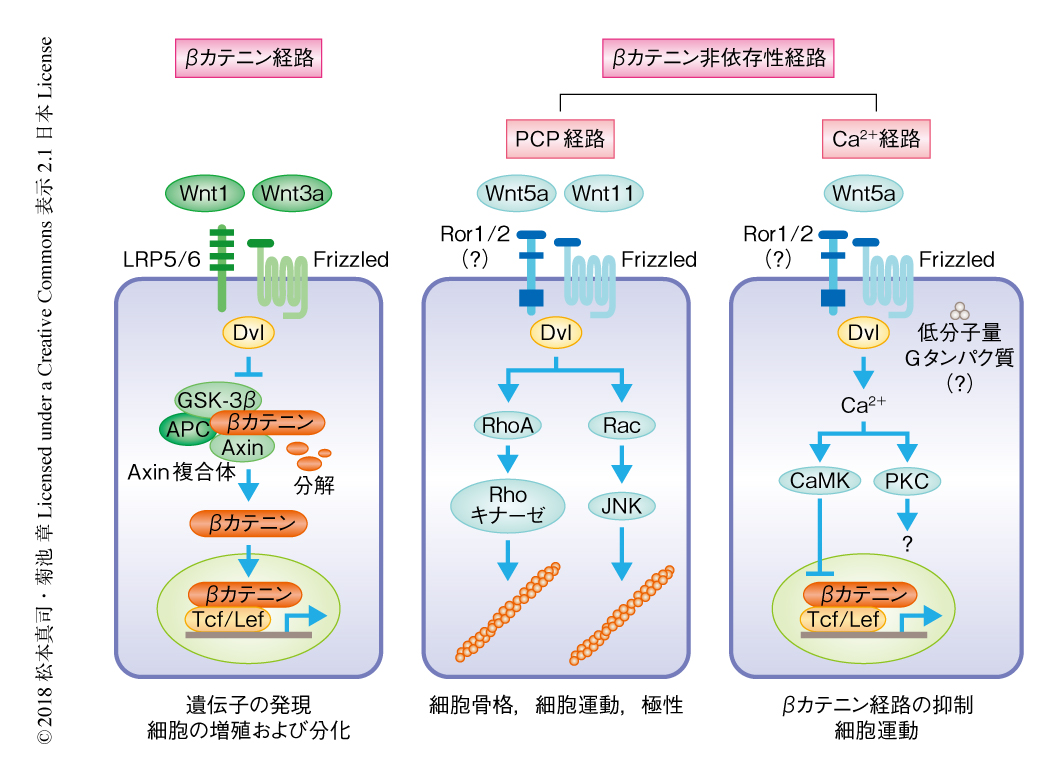
Wntは分子量が約4万の分泌性糖タンパク質で,ショウジョウバエから哺乳動物にいたるまで種をこえて保存され,初期発生や形態形成,また,出生ののちの増殖,分化,細胞運動,極性などを制御する2).Wntシグナル伝達経路には,βカテニンを介して遺伝子の発現を制御するβカテニン経路と,βカテニン経路とは独立し主として細胞骨格や細胞運動などを制御するβカテニン非依存性経路が存在する3)(図1).これまでに,リガンドとしてのWntはヒトおよびマウスにおいて19種類が同定されている2).また,Wntの受容体としては,7回膜貫通型のFrizzled(Fz1~Fz10の10種類)にくわえ,1回膜貫通型のLRP5,LRP6,Ror2,Rykが存在する3).少なくとも,βカテニン経路の活性化には1種類のFrizzledとLRP5あるいはLRP6が共役受容体として機能する.一方,Wnt5aはFrizzled,Ror1あるいはRor2と三者複合体を形成することによりβカテニン非依存性経路を活性化する4).
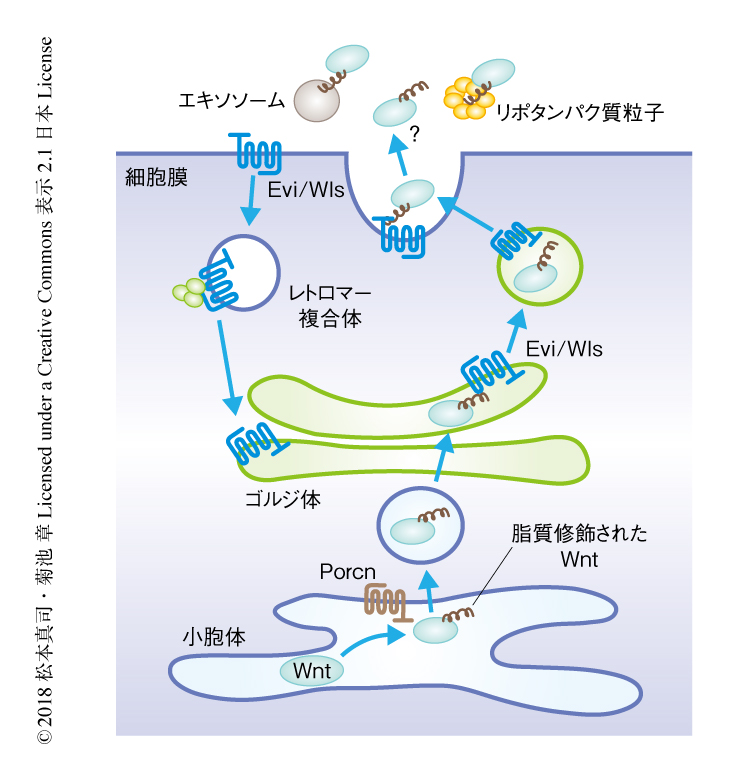
Wntタンパク質の翻訳後修飾と分泌:すべてのWntは合成の過程において小胞体に局在するアシル転移酵素Porcnによりパルミトイル化される.脂質修飾をうけたWntは,そののち,7回膜貫通型ソーティング受容体であるEvi/Wlsと結合し,ゴルジ体を介してWlsに依存的に細胞外へと分泌される.Wntの分泌にともない細胞膜に移行するWlsは,レトロマー複合体による逆行性輸送によりゴルジ体にもどされる5).Porcnによる脂質修飾はWntの分泌のみならず,受容体との結合や疎水的な細胞膜との親和性にも寄与する.ほとんどの場合,Wntはその疎水性から非常に近い距離にある細胞に対し作用すると考えられているが,分泌されたWntが標的になる細胞にまで輸送される機構については,直接的な分泌,エキソソーム,リポタンパク質粒子による輸送などが示唆されているものの,いまだ判然としない(図2).
βカテニン経路:βカテニンはカドヘリン結合タンパク質として同定され,細胞接着に重要なはたらきをもつと同時に,Wntシグナルのメディエーターとしても機能する.Wntによる刺激のない状態においては,細胞質のβカテニンがAxin,APC,GSK-3βからなるAxin複合体によるリン酸化と,それにともなうβ-TrCPによるユビキチン化をうけ,最終的にはプロテアソームにより分解されるため,βカテニンの量は低いレベルに保たれる3).Wnt3aやWnt1などが受容体であるFrizzledあるいは共役受容体であるLRP5,LRP6と結合すると,Dvlが細胞膜へとリクルートされる結果,Axin複合体の機能が抑制され,βカテニンは安定化して核へと移行したのち,転写因子であるTcf/Lefと結合し,Cyclin D1遺伝子やc-Myc遺伝子などの発現を促進することにより細胞の増殖や分化を制御する.
βカテニン非依存性経路:βカテニン非依存性経路には,平面内細胞極性(planar cell polarity:PCP)を制御するPCP経路と,細胞内におけるCa2+の動員を促進するCa2+経路の,少なくとも2種類が存在する3).Wnt5aやWnt11などが受容体であるFrizzledあるいは共役受容体であるRor1,Ror2,Rykと結合すると,細胞において多様なβカテニン非依存性経路が活性化される.第1の経路であるPCP経路は,ショウジョウバエの翅の表面において1層の上皮細胞から形成される翅毛の配向を決定するシグナル伝達経路として見い出された.PCP経路はFrizzledあるいはDvlを介してRhoファミリーの低分子量Gタンパク質を活性化し,さらに,RhoキナーゼやJNKの活性化により細胞骨格を制御して細胞極性や細胞運動,また,遺伝子の発現を促進する.第2の経路であるCa2+経路については,Wntが細胞内におけるCa2+の動員をひき起こし,CaMKおよびPKCを活性化することも報告されている.生理的な意義は不明であるが,βカテニン非依存性経路はβカテニン経路に対し拮抗することが示されている6).このように,PCP経路とCa2+経路のほかにもβカテニンを介さないシグナル伝達経路の存在が見い出されており,βカテニン非依存性経路は多様性が高いと考えられる7).
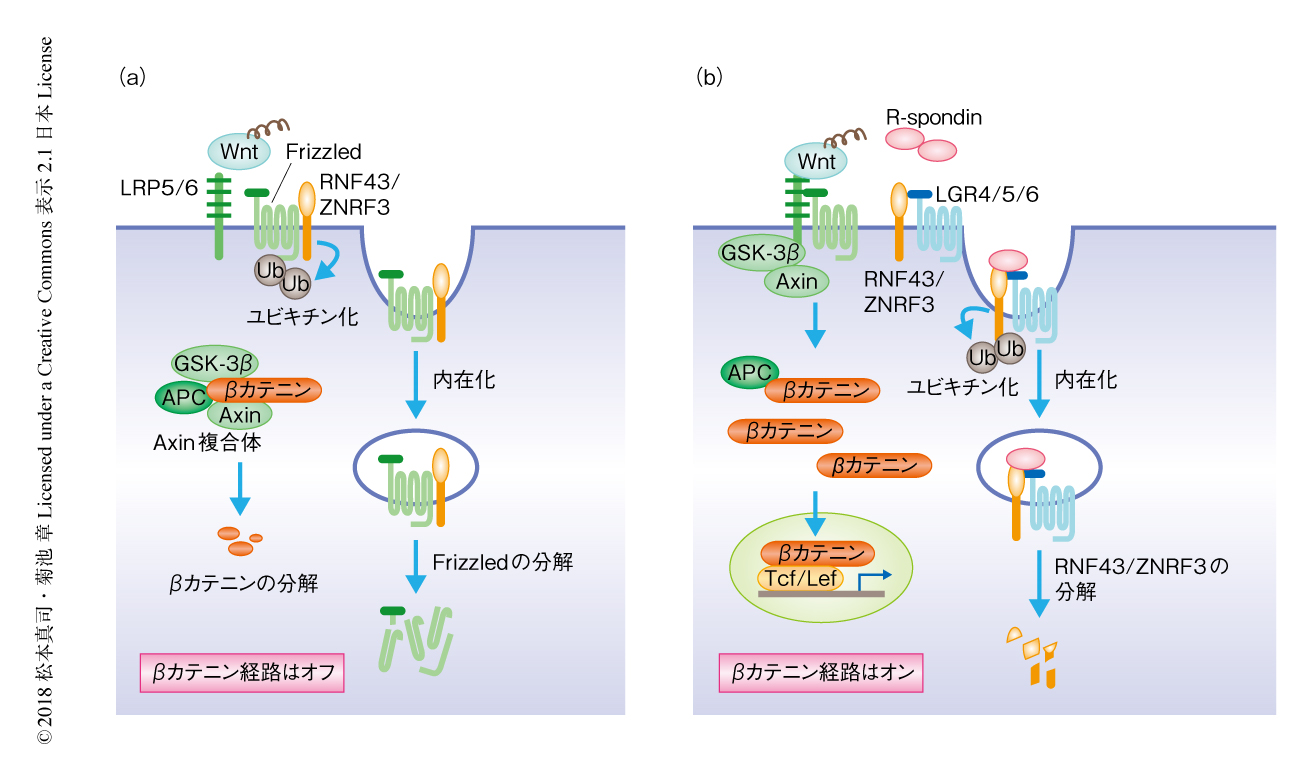
R-spondin経路:最近になり,WntおよびWnt受容体にくわえ,βカテニン経路を強力に増強する第3のタンパク質としてR-spondin(Rspo1~Rspo4の4種類)が同定され,ヒトのがんとのかかわりも報告されている.R-spondinの刺激のない状態においては,膜貫通型のユビキチンリガーゼであるRNF43/ZNRF3は細胞膜のFrizzledをユビキチン化し,Frizzledはリソソームにおいて分解される.R-spondinが細胞膜においてLGR4,LGR5,LGR6およびRNF43/ZNRF3と結合すると,RNF43/ZNRF3が自己ユビキチン化により細胞膜から除去され,その結果,Frizzledが細胞膜において安定化しβカテニン経路を増強する8)(図3).R-spondinシグナルはFrizzledの安定化をひき起こすことから,βカテニン経路にくわえβカテニン非依存性経路も活性化すると考えられているが,その生理的な意義については十分に解明されていない.
このように,Wntは複数の細胞内シグナル伝達機構を活性化することにより,多彩な細胞応答を制御する.したがって,Wntシグナル伝達経路に異常が生じると,がんをはじめとして種々の疾患がひき起こされる9).
近年,組織幹細胞をin vitroにおいて培養するオルガノイド培養技術が急速に発展し,Wntシグナルが腸管,胃,肝臓や膵臓といった多くの上皮組織幹細胞の増殖および維持に重要な役割をはたすことが明らかにされ,上皮を発生の母地とする多くのがんにWntシグナルが関与する可能性が示唆されている.実際,The Cancer Genome Atlas(TCGA)やInternational Cancer Genome Consortium(ICGC)といった大規模ながんゲノムプロジェクトにより,DNAメチル化や遺伝子あるいはタンパク質の発現の異常に関する網羅的なデータが公開され,Wntシグナル伝達経路の構成タンパク質の異常が多様ながん種において認められることが明らかにされている10).
APC遺伝子の異常:APC遺伝子は家族性大腸腺腫症の原因遺伝子として同定された11).APCタンパク質は約2800個のアミノ酸残基からなり,Axinおよびβカテニンと直接に結合してβカテニンの分解を促進する.APC遺伝子の異常は家族性大腸腺腫症にくわえ大腸がんにおいても約80%の症例で見い出されている.家族性大腸腺腫症や大腸がんにおけるAPC遺伝子の異常の大部分は,途中に終止コドンが生じAPCのC末端側の半分が欠損するものである.この変異APCはβカテニンとの結合能は保たれているが,Axinとは結合できない.したがって,βカテニンが効率よくリン酸化されないため,βカテニンの分解能が低下し蓄積する.最近になり,ヒトの大腸オルガノイドの培養モデルにおいて,CRISPR-Cas9法によるゲノム編集を用いてAPC遺伝子を含む複数の遺伝子に変異を導入することにより,多段階の発がんがin vitroにおいて再現され,さらに実際に大腸がんがひき起こされることが示され,発がんの過程におけるAPC遺伝子の異常の重要性があらためて確認された12).
βカテニン遺伝子の異常:大腸がんの約15%の症例でβカテニン遺伝子の異常が認められ,その変異はエキソン3に集中している11).この領域にはCK1αおよびGSK-3βによりリン酸化されるアミノ酸配列,および,β-TrCPによるユビキチン化の認識配列が存在する.これらのアミノ酸残基の変異,あるいは,エキソン3の完全欠損あるいは部分欠損により,βカテニンはCK1αやGSK-3βによりリン酸化されなくなる,あるいは,ユビキチン化をうけなくなり,変異βカテニンは細胞質や核に蓄積する.βカテニン遺伝子の異常は,肝臓がん,副腎皮質がん,胃がん,前立腺がん,小児肝がんである肝芽腫などにおいて高い頻度で認められる.
Axin遺伝子の異常:ヒトにはAxin1遺伝子およびAxin2遺伝子(ラットAxil遺伝子,マウスconductin遺伝子)の2つのAxin遺伝子が存在し,肝臓がんにおいてAxin1遺伝子,大腸がんにおいてAxin2遺伝子の異常が報告されている13,14).Axinの異常によりAPC,βカテニン,GSK-3βとの複合体が形成されなくなり,βカテニンのリン酸化やユビキチン化が抑制され,その結果,βカテニンが蓄積すると考えられている.
R-spondin経路における遺伝子の異常:最近になり,RNF43遺伝子の機能欠失変異が膵がん15) および大腸がん16) において,また,ZNRF3遺伝子の変異が副腎皮質がん17) において,はじめて報告された.さらに,Rspo2遺伝子あるいはRspo3遺伝子の融合が約10%の大腸がんにおいて認められている18).大腸がんにおいて,RNF43遺伝子の変異あるいはR-spondin遺伝子の融合とAPC遺伝子の変異は相互排他的な関係にあることから,Wntシグナル伝達経路の活性化および腫瘍の形成に関与すると考えられる.一方で,RNF43/ZNRF3遺伝子に変異をもつ膵がんおよび大腸がんはPorcnの阻害剤に感受性を示すことから,細胞の増殖がWntの分泌に強く依存することが明らかにされた19,20).
Wnt5aのがんの悪性化作用:膵がんや悪性黒色腫をはじめとするヒトの多くのがんにおいて,βカテニン非依存性経路を活性化するWnt5aは細胞運動や浸潤能を促進し,腫瘍の進展に関与する6).非小細胞肺がんにおいてはWnt5a遺伝子の過剰な発現と腫瘍の増殖および間質における血管の新生とのあいだに正の相関が認められる21).胃がんにおいても,Wnt5a遺伝子は約30%の症例において過剰に発現しており,悪性度の高いスキルス型においてWnt5a陽性の症例が有意に多い22).さらに,Wnt5a陽性例の術後5年の生存率は陰性例に比べ有意に低い.また,Wnt5a遺伝子が高発現している前立腺がんの症例では病理組織学的な悪性度が高く,術後の再発率も高い23).したがって,Wnt5aはがん種に応じてがんの悪性化を促進する作用をもつことから,ある種のがんにおいては診断マーカーや予後の判定指標,あるいは,治療の標的になる可能性がある.
Wnt5aのがんの抑制作用:これに対し,Wnt5aはβカテニン経路を抑制するはたらきがあることから,がんの抑制作用をもつとも考えられている6).たとえば,甲状腺がんや卵巣がんの細胞株においてWnt5aは細胞の増殖を抑制する24,25).これらの結果と一致して,Wnt5aのヘテロノックアウトマウスを長期間にわたり飼育すると,24カ月以内に約20%のマウスにおいてBリンパ腫あるいは慢性骨髄性白血病が自然発症する26).ヒトの白血病においても,Wnt5a遺伝子の発現が抑制される症例が認められる.
βカテニン経路を制御するAxin複合体の構成タンパク質の遺伝子の異常とがんとの関係が明らかにされたことから,これまで,βカテニン経路を直接に阻害する抗がん剤の開発が進められてきたが,いまだ実用化にはいたっていない.そこで,Wntシグナル伝達経路に関連する創薬の標的になりうる新規の経路の探索が進められている(図4).
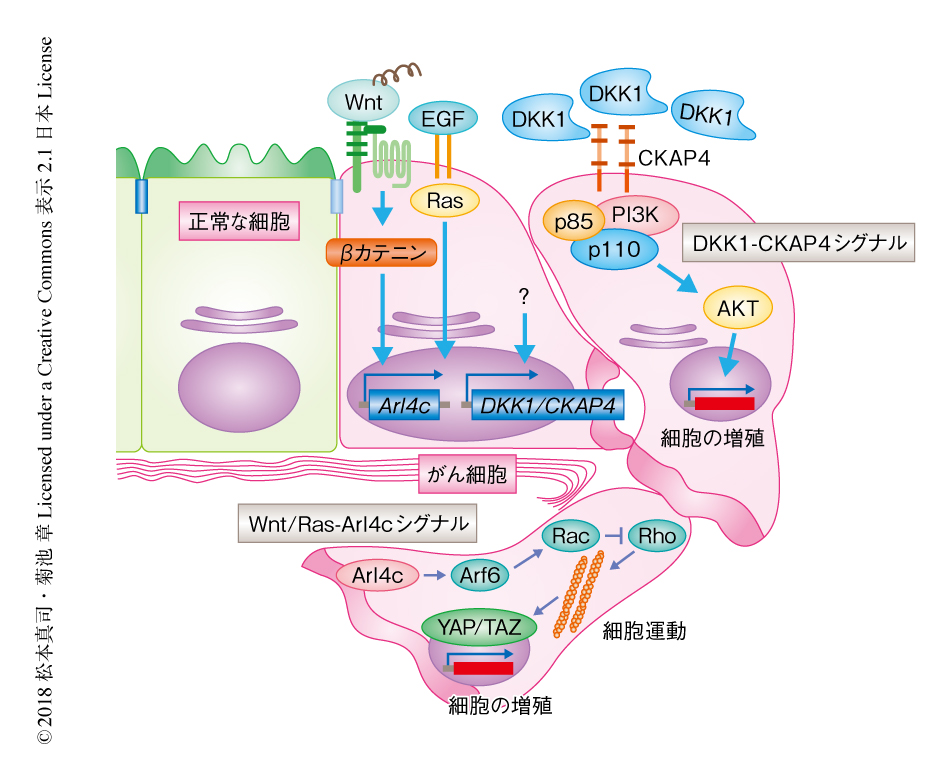
DKK1によるがん細胞の増殖能の促進の分子機構:DKK1は,LRP6と結合しβカテニン経路を阻害する分泌性タンパク質で,動物の発生に必須である27).出生ののちの組織における発現はきわめて低いが,がん組織において特異的な高発現が認められる.その作用機構から,sFRP1やsFRP2と同様に,DKK1はがんの抑制能をもつと考えられていたが,DKK1遺伝子が肺がんや食道がんにおいて高発現することや,抗DKK1抗体が肺がん細胞株の増殖を抑制することから,DKK1がWntシグナルの阻害とは関係なく,細胞の増殖を促進する可能性も示唆されていた28-30).DKK1結合タンパク質の探索の結果,細胞膜1回膜貫通II型膜タンパク質であるCKAP4がDKK1の新規の受容体であることが判明した.DKK1-CKAP4シグナル軸は,PI3KおよびAKTを活性化することにより細胞の増殖を促進する31).DKK1遺伝子およびCKAP4遺伝子は膵がん,肺腺がん,肺扁平上皮がん,食道扁平上皮がんの約40~60%の症例においてがん組織に特異的に発現しており,DKK1遺伝子およびCKAP4遺伝子が発現している症例は,DKK1遺伝子およびCKAP4遺伝子がともに発現していない症例,および,いずれか一方のみが発現している症例に比べ,予後が有意に不良であった31,32).くわえて,DKK1と同様にがんの促進効果が認められながらこれまで受容体が不明であったDKK3も,食道がんの約50%の症例においてp53ファミリーの転写因子であるp63(TP63)に依存的に過剰に発現し,CKAP4を介して細胞の増殖および腫瘍の形成を促進することが明らかにされている33).さらに,抗CKAP4抗体が,DKK1遺伝子あるいはDKK3遺伝子を発現する膵がん細胞株,肺がん細胞株,食道がん細胞株のin vivoにおける腫瘍の形成を抑制することから,CKAP4はある種のがんにおいて新規の診断マーカーや治療の標的になると考えられる.
Arl4cによるがん細胞の運動および増殖能の促進の分子機構:ヒトの多くのがんにおいてドライバーシグナルとして機能するEGF-RasシグナルおよびWnt-βカテニンシグナルの共通の標的遺伝子として,Arl4c遺伝子が同定された34).正常な上皮細胞における管状構造の形成の過程は,がん細胞が運動能および増殖能を獲得して間質へと侵入していく過程と類似している.Wnt-βカテニンシグナルとEGF-Rasシグナルが同時協調的に活性化するとArl4c遺伝子が発現し,上皮細胞の形態変化と活発な増殖を介して管状構造の形成をひき起こす.Arl4cは別種の低分子量Gタンパク質であるArf6,Rac,Rhoの活性を制御し,その結果,YAP/YAZが核へと移行することにより細胞の増殖を促進する34).また,Arl4c遺伝子は大腸がん,肺腺がん,肺扁平上皮がん,舌扁平上皮がんの約50~80%の症例において腫瘍部に特異的に高発現しており,細胞の運動能,浸潤能,増殖能を促進する35,36).くわえて,最近では,Arl4c遺伝子の発現が胃がんの腹膜播種を促進することも報告されている37).Arl4c遺伝子は大腸がん細胞株,肺がん細胞株,肝がん細胞株,胃がん細胞株において,Wnt-βカテニンシグナルあるいはEGF-Rasシグナルに依存して過剰に発現する.大腸がん細胞株や肝がん細胞株のin vivoにおける腫瘍の形成は,Arl4cに対するsiRNAやアンチセンス核酸の投与により抑制されることから,新たなWnt関連タンパク質として治療の標的になる可能性がある38).
APCやβカテニンといったAxin複合体の構成タンパク質の遺伝子変異の頻度の高さから,Wntシグナルを標的にした初期の阻害剤の開発は,おもにβカテニン-TCF複合体を標的にして精力的に進められてきた.一般に,核内転写因子に対する阻害剤の開発はむずかしいとされていて,βカテニン-TCF複合体に対しても,いまだ治療に用いられるような薬剤は存在しない.一方で,近年,Wntの分泌経路が詳細に明らかにされたことで,WntやWnt受容体のレベルでの阻害剤の開発が進められている.以下,現在,臨床試験が行われている主要なWntシグナル阻害剤について解説する(図5).
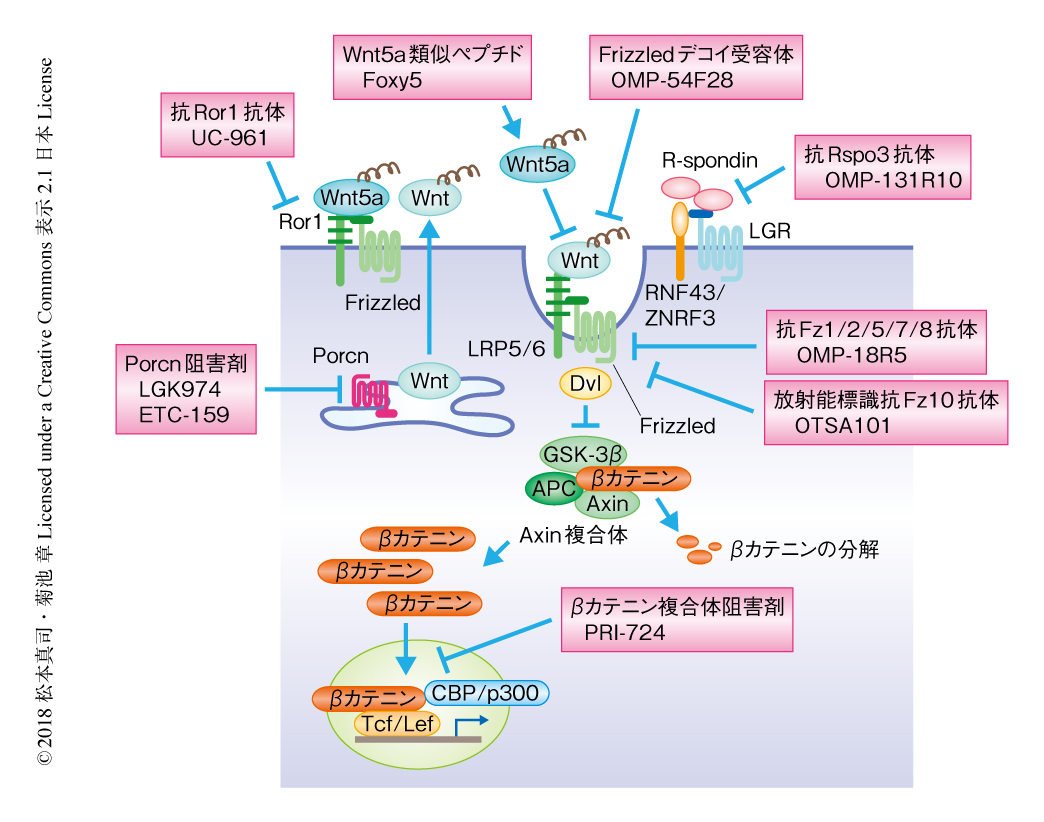
PorcnあるいはWntの分泌阻害剤:さきに述べたように,Wntの細胞外への分泌はPorcnによる脂質修飾に依存することが明らかにされ,Porcnの阻害剤の開発が進んでいる.これまで,IWP-2,WNT-C59,LGK974(WNT974),ETC-159(ETC-1922159)が開発され,RNF43遺伝子に変異をもつ膵がん39),RNF43遺伝子あるいはR-spondin遺伝子の融合をもつ大腸がん40) に対する抗腫瘍効果が確認されている.このうち,LGK974およびETC-159は難治性の大腸がんに対し第1相の臨床試験が行われている.
細胞外のWnt関連リガンドあるいはWnt関連受容体に対する阻害剤:近年,特定のWnt関連リガンドやWnt関連受容体の異常とがんの関連も報告され,多くの阻害剤が開発されている.
OMP-54F28はFz8のシステインリッチドメインとヒト免疫グロブリンGのFc領域との融合タンパク質であり,Wntに対するデコイ受容体として機能しβカテニン経路を阻害する41).OMP-54F28はヒトの肝がんおよび卵巣がんのマウスへの異種移植による腫瘍の形成を抑制することが確認されており,現在,既存の化学療法との併用により肝がん,卵巣がん,膵がんに対し第1相の臨床試験が行われている.
OMP-131R10はRspo3に対するモノクローナル抗体で,Rspo3遺伝子の融合をもつ大腸がん,あるいは,Rspo3遺伝子を過剰に発現する非小細胞肺がんの異種移植による腫瘍の形成を抑制する.現在,転移性の大腸がんに対して第1相の臨床試験が行われている.
OMP-18R5はFz7に対するモノクローナル抗体として作製されたが,くわえて,Fz1,Fz2,Fz5,Fz8に対する認識能をもち,βカテニン経路を阻害することにより種々のヒトのがん細胞の異種移植による腫瘍の形成を抑制する42).現在,膵がん,転移性乳がん,非小細胞肺がんに対し第1相の臨床試験が行われている.
OTSA101は放射性同位元素により標識したFz10に対するモノクローナル抗体で,Fz10が滑膜肉腫において特異的に過剰に発現することから,現在,滑膜肉腫に対し第1相の臨床試験が行われている43).
βカテニンを含む複合体に対する阻害剤:PRI-724はβカテニンと転写共役因子であるCBPとの結合を阻害する低分子化合物で,βカテニンに依存的な遺伝子の発現を阻害する44).現在,βカテニンを含む複合体に対する阻害剤としては唯一,急性骨髄性白血病,慢性骨髄性白血病,大腸がん,膵がんに対し第1相および第2相の臨床試験が行われている.
βカテニン非依存性経路を標的にした抗がん剤:Cirmtuzumab(UC-961)はRor1に対するモノクローナル抗体で,Wnt5a-Ror1シグナル伝達経路を阻害することにより,Ror1遺伝子を過剰に発現する慢性リンパ性白血病細胞の増殖を抑制する45,46).現在,慢性リンパ性白血病に対し第1相の臨床試験が行われている.Foxy5はWnt5aの類似ペプチドで,Wnt5aシグナルのアゴニストとして作用する.さきに述べたように,Wnt5aはある種のがんにおいて抑制的に作用するが,実際,マウスモデルにおいてFoxy5は乳がん細胞の肺および肝転移を抑制する47).また,Wnt5a遺伝子の低発現は乳がん,大腸がん,前立腺がんにおいて予後不良と相関することから48-50),現在,これらのがんに対して第1相の臨床試験が行われている.しかし,Wnt5aはβカテニン非依存性経路を介してがんに促進的に作用することもあるので,現状では,Wnt5aのアゴニストが抗腫瘍効果を発揮するがんの明確な識別は困難である.現在,これらを識別するためのマーカー遺伝子の探索が試みられている51).
35年以上におよぶWntの研究の結果,Wntシグナル伝達経路の異常がヒトの多様ながんに関与することが明らかにされてきた.しかし,Wntシグナルが毛包や腸管をはじめとする多くの正常な組織幹細胞の維持,増殖,分化にきわめて重要な役割をもつことから,Wntシグナルを標的にした阻害剤にはつねに深刻な副作用が懸念される.そのため,主要なWntシグナル伝達経路の構成タンパク質にくわえ,がんに特異的に機能する新たなWnt関連タンパク質やシグナル伝達経路を同定する試みは,今後,ますます重要になると考えられる.また,既存の分子標的薬や抗腫瘍薬と組み合わせてWntシグナルを阻害する試みも,副作用の回避につながるかもしれない.
最近になり,Wntシグナルはがん細胞そのものにくわえ,周囲のがん微小環境に対しても多様なはたらきをもつことが示されている.とくに,腫瘍免疫に対する作用は,今後,ますます注目されるだろう.実際,Wnt-βカテニンシグナルの活性化が腫瘍においてCD8陽性T細胞の排除にはたらくなど,抗腫瘍免疫を抑制する可能性が示唆されており52),Wntシグナル伝達経路の阻害剤とがん免疫療法との併用による治療効果の改善も期待される.
略歴:2009年 広島大学大学院医歯薬学総合研究科博士課程 修了,同年 大阪大学大学院医学系研究科 特任研究員を経て,2012年より同 特任助教(現 助教).
研究テーマ:発生の過程から疾患まで多岐にわたるWntシグナルの生理的な意義.
抱負:数多くある増殖因子のなかで,Wntシグナルが具体的になぜ,どのように大事なのか,生理と病理の両面からその本質にせまりたい.
菊池 章(Akira Kikuchi)
大阪大学大学院医学系研究科 教授.
研究室URL:http://www.med.osaka-u.ac.jp/pub/molbiobc/
© 2018 松本真司・菊池 章 Licensed under CC 表示 2.1 日本
(大阪大学大学院医学系研究科 分子病態生化学)
email:松本真司,菊池 章
領域融合レビュー, 7, e009 (2018) DOI: 10.7875/leading.author.7.e009
Shinji Matsumoto & Akira Kikuchi: Wnt signaling pathways in cancers: key targets and implications in cancer therapy.
要 約
Wntは分泌性糖タンパク質であり,Wntシグナル伝達経路を活性化することにより,増殖,分化,細胞運動,極性など,多岐にわたる細胞の応答を制御する.Wntシグナルは動物の胎生期における発生の過程に必須であり,種々の臓器の形成を制御する.出生ののちには臓器の恒常性の維持に関与し,再生や修復の過程において活性化される.一方,大腸がんをはじめとするヒトの種々のがんにおいて,Wntシグナル伝達経路の異常な活性化の関与について報告されている.これまで,Wntシグナル伝達経路の遮断を目的とした抗がん剤の開発が試みられてきたものの,医薬品として実用化された例はない.しかし,近年のヒトのがんゲノムの網羅的な解析やオルガノイド培養技術とあいまって,新たながんシグナルタンパク質の発見および分子標的治療薬の開発が期待されている.
はじめに
1982年,マウスの乳がんの原因遺伝子としてInt-1遺伝子がクローニングされ,ショウジョウバエにおけるInt-1遺伝子のホモログがWingless遺伝子という分節の形成に重要な役割をはたす遺伝子であったことから,Int-1はWnt-1(Wingless + Int-1)と名づけられた1).それから35年以上がたち,腫瘍医学,発生生物学,幹細胞生物学においても解析が進み,Wntの研究領域は大きく展開してきた.ヒトのがんにおけるWntシグナルの重要性は,おもに大腸がんにおいて明らかにされてきた.しかし,近年のヒトのがんゲノムの網羅的な解析や,幹細胞の研究において発展したオルガノイド培養技術の応用などにより,Wntシグナル伝達経路の構成タンパク質およびWnt関連タンパク質の異常がヒトの多様ながんに関与することが明らかにされてきた.一方で,これまで,Wntシグナル伝達経路の遮断を目的とした抗がん剤の開発が試みられてきたものの,効果と安全性の問題から医薬品として実用化された例はない.このレビューにおいては,これまでに明らかにされてきたWntシグナルの活性化の機構に関する知見から,ヒトのがんにおけるWntシグナル伝達経路の構成タンパク質の異常,また,現在,臨床試験が進められているWnt関連タンパク質を標的にした抗がん剤の開発について解説する.
1.Wntシグナル伝達経路の概要
Wntは分子量が約4万の分泌性糖タンパク質で,ショウジョウバエから哺乳動物にいたるまで種をこえて保存され,初期発生や形態形成,また,出生ののちの増殖,分化,細胞運動,極性などを制御する2).Wntシグナル伝達経路には,βカテニンを介して遺伝子の発現を制御するβカテニン経路と,βカテニン経路とは独立し主として細胞骨格や細胞運動などを制御するβカテニン非依存性経路が存在する3)(図1).これまでに,リガンドとしてのWntはヒトおよびマウスにおいて19種類が同定されている2).また,Wntの受容体としては,7回膜貫通型のFrizzled(Fz1~Fz10の10種類)にくわえ,1回膜貫通型のLRP5,LRP6,Ror2,Rykが存在する3).少なくとも,βカテニン経路の活性化には1種類のFrizzledとLRP5あるいはLRP6が共役受容体として機能する.一方,Wnt5aはFrizzled,Ror1あるいはRor2と三者複合体を形成することによりβカテニン非依存性経路を活性化する4).
Wntタンパク質の翻訳後修飾と分泌:すべてのWntは合成の過程において小胞体に局在するアシル転移酵素Porcnによりパルミトイル化される.脂質修飾をうけたWntは,そののち,7回膜貫通型ソーティング受容体であるEvi/Wlsと結合し,ゴルジ体を介してWlsに依存的に細胞外へと分泌される.Wntの分泌にともない細胞膜に移行するWlsは,レトロマー複合体による逆行性輸送によりゴルジ体にもどされる5).Porcnによる脂質修飾はWntの分泌のみならず,受容体との結合や疎水的な細胞膜との親和性にも寄与する.ほとんどの場合,Wntはその疎水性から非常に近い距離にある細胞に対し作用すると考えられているが,分泌されたWntが標的になる細胞にまで輸送される機構については,直接的な分泌,エキソソーム,リポタンパク質粒子による輸送などが示唆されているものの,いまだ判然としない(図2).
βカテニン経路:βカテニンはカドヘリン結合タンパク質として同定され,細胞接着に重要なはたらきをもつと同時に,Wntシグナルのメディエーターとしても機能する.Wntによる刺激のない状態においては,細胞質のβカテニンがAxin,APC,GSK-3βからなるAxin複合体によるリン酸化と,それにともなうβ-TrCPによるユビキチン化をうけ,最終的にはプロテアソームにより分解されるため,βカテニンの量は低いレベルに保たれる3).Wnt3aやWnt1などが受容体であるFrizzledあるいは共役受容体であるLRP5,LRP6と結合すると,Dvlが細胞膜へとリクルートされる結果,Axin複合体の機能が抑制され,βカテニンは安定化して核へと移行したのち,転写因子であるTcf/Lefと結合し,Cyclin D1遺伝子やc-Myc遺伝子などの発現を促進することにより細胞の増殖や分化を制御する.
βカテニン非依存性経路:βカテニン非依存性経路には,平面内細胞極性(planar cell polarity:PCP)を制御するPCP経路と,細胞内におけるCa2+の動員を促進するCa2+経路の,少なくとも2種類が存在する3).Wnt5aやWnt11などが受容体であるFrizzledあるいは共役受容体であるRor1,Ror2,Rykと結合すると,細胞において多様なβカテニン非依存性経路が活性化される.第1の経路であるPCP経路は,ショウジョウバエの翅の表面において1層の上皮細胞から形成される翅毛の配向を決定するシグナル伝達経路として見い出された.PCP経路はFrizzledあるいはDvlを介してRhoファミリーの低分子量Gタンパク質を活性化し,さらに,RhoキナーゼやJNKの活性化により細胞骨格を制御して細胞極性や細胞運動,また,遺伝子の発現を促進する.第2の経路であるCa2+経路については,Wntが細胞内におけるCa2+の動員をひき起こし,CaMKおよびPKCを活性化することも報告されている.生理的な意義は不明であるが,βカテニン非依存性経路はβカテニン経路に対し拮抗することが示されている6).このように,PCP経路とCa2+経路のほかにもβカテニンを介さないシグナル伝達経路の存在が見い出されており,βカテニン非依存性経路は多様性が高いと考えられる7).
R-spondin経路:最近になり,WntおよびWnt受容体にくわえ,βカテニン経路を強力に増強する第3のタンパク質としてR-spondin(Rspo1~Rspo4の4種類)が同定され,ヒトのがんとのかかわりも報告されている.R-spondinの刺激のない状態においては,膜貫通型のユビキチンリガーゼであるRNF43/ZNRF3は細胞膜のFrizzledをユビキチン化し,Frizzledはリソソームにおいて分解される.R-spondinが細胞膜においてLGR4,LGR5,LGR6およびRNF43/ZNRF3と結合すると,RNF43/ZNRF3が自己ユビキチン化により細胞膜から除去され,その結果,Frizzledが細胞膜において安定化しβカテニン経路を増強する8)(図3).R-spondinシグナルはFrizzledの安定化をひき起こすことから,βカテニン経路にくわえβカテニン非依存性経路も活性化すると考えられているが,その生理的な意義については十分に解明されていない.
このように,Wntは複数の細胞内シグナル伝達機構を活性化することにより,多彩な細胞応答を制御する.したがって,Wntシグナル伝達経路に異常が生じると,がんをはじめとして種々の疾患がひき起こされる9).
2.βカテニン経路の異常とがん
近年,組織幹細胞をin vitroにおいて培養するオルガノイド培養技術が急速に発展し,Wntシグナルが腸管,胃,肝臓や膵臓といった多くの上皮組織幹細胞の増殖および維持に重要な役割をはたすことが明らかにされ,上皮を発生の母地とする多くのがんにWntシグナルが関与する可能性が示唆されている.実際,The Cancer Genome Atlas(TCGA)やInternational Cancer Genome Consortium(ICGC)といった大規模ながんゲノムプロジェクトにより,DNAメチル化や遺伝子あるいはタンパク質の発現の異常に関する網羅的なデータが公開され,Wntシグナル伝達経路の構成タンパク質の異常が多様ながん種において認められることが明らかにされている10).
APC遺伝子の異常:APC遺伝子は家族性大腸腺腫症の原因遺伝子として同定された11).APCタンパク質は約2800個のアミノ酸残基からなり,Axinおよびβカテニンと直接に結合してβカテニンの分解を促進する.APC遺伝子の異常は家族性大腸腺腫症にくわえ大腸がんにおいても約80%の症例で見い出されている.家族性大腸腺腫症や大腸がんにおけるAPC遺伝子の異常の大部分は,途中に終止コドンが生じAPCのC末端側の半分が欠損するものである.この変異APCはβカテニンとの結合能は保たれているが,Axinとは結合できない.したがって,βカテニンが効率よくリン酸化されないため,βカテニンの分解能が低下し蓄積する.最近になり,ヒトの大腸オルガノイドの培養モデルにおいて,CRISPR-Cas9法によるゲノム編集を用いてAPC遺伝子を含む複数の遺伝子に変異を導入することにより,多段階の発がんがin vitroにおいて再現され,さらに実際に大腸がんがひき起こされることが示され,発がんの過程におけるAPC遺伝子の異常の重要性があらためて確認された12).
βカテニン遺伝子の異常:大腸がんの約15%の症例でβカテニン遺伝子の異常が認められ,その変異はエキソン3に集中している11).この領域にはCK1αおよびGSK-3βによりリン酸化されるアミノ酸配列,および,β-TrCPによるユビキチン化の認識配列が存在する.これらのアミノ酸残基の変異,あるいは,エキソン3の完全欠損あるいは部分欠損により,βカテニンはCK1αやGSK-3βによりリン酸化されなくなる,あるいは,ユビキチン化をうけなくなり,変異βカテニンは細胞質や核に蓄積する.βカテニン遺伝子の異常は,肝臓がん,副腎皮質がん,胃がん,前立腺がん,小児肝がんである肝芽腫などにおいて高い頻度で認められる.
Axin遺伝子の異常:ヒトにはAxin1遺伝子およびAxin2遺伝子(ラットAxil遺伝子,マウスconductin遺伝子)の2つのAxin遺伝子が存在し,肝臓がんにおいてAxin1遺伝子,大腸がんにおいてAxin2遺伝子の異常が報告されている13,14).Axinの異常によりAPC,βカテニン,GSK-3βとの複合体が形成されなくなり,βカテニンのリン酸化やユビキチン化が抑制され,その結果,βカテニンが蓄積すると考えられている.
R-spondin経路における遺伝子の異常:最近になり,RNF43遺伝子の機能欠失変異が膵がん15) および大腸がん16) において,また,ZNRF3遺伝子の変異が副腎皮質がん17) において,はじめて報告された.さらに,Rspo2遺伝子あるいはRspo3遺伝子の融合が約10%の大腸がんにおいて認められている18).大腸がんにおいて,RNF43遺伝子の変異あるいはR-spondin遺伝子の融合とAPC遺伝子の変異は相互排他的な関係にあることから,Wntシグナル伝達経路の活性化および腫瘍の形成に関与すると考えられる.一方で,RNF43/ZNRF3遺伝子に変異をもつ膵がんおよび大腸がんはPorcnの阻害剤に感受性を示すことから,細胞の増殖がWntの分泌に強く依存することが明らかにされた19,20).
3.βカテニン非依存性経路の異常とがん
Wnt5aのがんの悪性化作用:膵がんや悪性黒色腫をはじめとするヒトの多くのがんにおいて,βカテニン非依存性経路を活性化するWnt5aは細胞運動や浸潤能を促進し,腫瘍の進展に関与する6).非小細胞肺がんにおいてはWnt5a遺伝子の過剰な発現と腫瘍の増殖および間質における血管の新生とのあいだに正の相関が認められる21).胃がんにおいても,Wnt5a遺伝子は約30%の症例において過剰に発現しており,悪性度の高いスキルス型においてWnt5a陽性の症例が有意に多い22).さらに,Wnt5a陽性例の術後5年の生存率は陰性例に比べ有意に低い.また,Wnt5a遺伝子が高発現している前立腺がんの症例では病理組織学的な悪性度が高く,術後の再発率も高い23).したがって,Wnt5aはがん種に応じてがんの悪性化を促進する作用をもつことから,ある種のがんにおいては診断マーカーや予後の判定指標,あるいは,治療の標的になる可能性がある.
Wnt5aのがんの抑制作用:これに対し,Wnt5aはβカテニン経路を抑制するはたらきがあることから,がんの抑制作用をもつとも考えられている6).たとえば,甲状腺がんや卵巣がんの細胞株においてWnt5aは細胞の増殖を抑制する24,25).これらの結果と一致して,Wnt5aのヘテロノックアウトマウスを長期間にわたり飼育すると,24カ月以内に約20%のマウスにおいてBリンパ腫あるいは慢性骨髄性白血病が自然発症する26).ヒトの白血病においても,Wnt5a遺伝子の発現が抑制される症例が認められる.
4.Wntシグナル関連タンパク質の発現の異常と新規のがんシグナル
βカテニン経路を制御するAxin複合体の構成タンパク質の遺伝子の異常とがんとの関係が明らかにされたことから,これまで,βカテニン経路を直接に阻害する抗がん剤の開発が進められてきたが,いまだ実用化にはいたっていない.そこで,Wntシグナル伝達経路に関連する創薬の標的になりうる新規の経路の探索が進められている(図4).
DKK1によるがん細胞の増殖能の促進の分子機構:DKK1は,LRP6と結合しβカテニン経路を阻害する分泌性タンパク質で,動物の発生に必須である27).出生ののちの組織における発現はきわめて低いが,がん組織において特異的な高発現が認められる.その作用機構から,sFRP1やsFRP2と同様に,DKK1はがんの抑制能をもつと考えられていたが,DKK1遺伝子が肺がんや食道がんにおいて高発現することや,抗DKK1抗体が肺がん細胞株の増殖を抑制することから,DKK1がWntシグナルの阻害とは関係なく,細胞の増殖を促進する可能性も示唆されていた28-30).DKK1結合タンパク質の探索の結果,細胞膜1回膜貫通II型膜タンパク質であるCKAP4がDKK1の新規の受容体であることが判明した.DKK1-CKAP4シグナル軸は,PI3KおよびAKTを活性化することにより細胞の増殖を促進する31).DKK1遺伝子およびCKAP4遺伝子は膵がん,肺腺がん,肺扁平上皮がん,食道扁平上皮がんの約40~60%の症例においてがん組織に特異的に発現しており,DKK1遺伝子およびCKAP4遺伝子が発現している症例は,DKK1遺伝子およびCKAP4遺伝子がともに発現していない症例,および,いずれか一方のみが発現している症例に比べ,予後が有意に不良であった31,32).くわえて,DKK1と同様にがんの促進効果が認められながらこれまで受容体が不明であったDKK3も,食道がんの約50%の症例においてp53ファミリーの転写因子であるp63(TP63)に依存的に過剰に発現し,CKAP4を介して細胞の増殖および腫瘍の形成を促進することが明らかにされている33).さらに,抗CKAP4抗体が,DKK1遺伝子あるいはDKK3遺伝子を発現する膵がん細胞株,肺がん細胞株,食道がん細胞株のin vivoにおける腫瘍の形成を抑制することから,CKAP4はある種のがんにおいて新規の診断マーカーや治療の標的になると考えられる.
Arl4cによるがん細胞の運動および増殖能の促進の分子機構:ヒトの多くのがんにおいてドライバーシグナルとして機能するEGF-RasシグナルおよびWnt-βカテニンシグナルの共通の標的遺伝子として,Arl4c遺伝子が同定された34).正常な上皮細胞における管状構造の形成の過程は,がん細胞が運動能および増殖能を獲得して間質へと侵入していく過程と類似している.Wnt-βカテニンシグナルとEGF-Rasシグナルが同時協調的に活性化するとArl4c遺伝子が発現し,上皮細胞の形態変化と活発な増殖を介して管状構造の形成をひき起こす.Arl4cは別種の低分子量Gタンパク質であるArf6,Rac,Rhoの活性を制御し,その結果,YAP/YAZが核へと移行することにより細胞の増殖を促進する34).また,Arl4c遺伝子は大腸がん,肺腺がん,肺扁平上皮がん,舌扁平上皮がんの約50~80%の症例において腫瘍部に特異的に高発現しており,細胞の運動能,浸潤能,増殖能を促進する35,36).くわえて,最近では,Arl4c遺伝子の発現が胃がんの腹膜播種を促進することも報告されている37).Arl4c遺伝子は大腸がん細胞株,肺がん細胞株,肝がん細胞株,胃がん細胞株において,Wnt-βカテニンシグナルあるいはEGF-Rasシグナルに依存して過剰に発現する.大腸がん細胞株や肝がん細胞株のin vivoにおける腫瘍の形成は,Arl4cに対するsiRNAやアンチセンス核酸の投与により抑制されることから,新たなWnt関連タンパク質として治療の標的になる可能性がある38).
5.Wntシグナル伝達経路を標的にした抗がん剤の開発
APCやβカテニンといったAxin複合体の構成タンパク質の遺伝子変異の頻度の高さから,Wntシグナルを標的にした初期の阻害剤の開発は,おもにβカテニン-TCF複合体を標的にして精力的に進められてきた.一般に,核内転写因子に対する阻害剤の開発はむずかしいとされていて,βカテニン-TCF複合体に対しても,いまだ治療に用いられるような薬剤は存在しない.一方で,近年,Wntの分泌経路が詳細に明らかにされたことで,WntやWnt受容体のレベルでの阻害剤の開発が進められている.以下,現在,臨床試験が行われている主要なWntシグナル阻害剤について解説する(図5).
PorcnあるいはWntの分泌阻害剤:さきに述べたように,Wntの細胞外への分泌はPorcnによる脂質修飾に依存することが明らかにされ,Porcnの阻害剤の開発が進んでいる.これまで,IWP-2,WNT-C59,LGK974(WNT974),ETC-159(ETC-1922159)が開発され,RNF43遺伝子に変異をもつ膵がん39),RNF43遺伝子あるいはR-spondin遺伝子の融合をもつ大腸がん40) に対する抗腫瘍効果が確認されている.このうち,LGK974およびETC-159は難治性の大腸がんに対し第1相の臨床試験が行われている.
細胞外のWnt関連リガンドあるいはWnt関連受容体に対する阻害剤:近年,特定のWnt関連リガンドやWnt関連受容体の異常とがんの関連も報告され,多くの阻害剤が開発されている.
OMP-54F28はFz8のシステインリッチドメインとヒト免疫グロブリンGのFc領域との融合タンパク質であり,Wntに対するデコイ受容体として機能しβカテニン経路を阻害する41).OMP-54F28はヒトの肝がんおよび卵巣がんのマウスへの異種移植による腫瘍の形成を抑制することが確認されており,現在,既存の化学療法との併用により肝がん,卵巣がん,膵がんに対し第1相の臨床試験が行われている.
OMP-131R10はRspo3に対するモノクローナル抗体で,Rspo3遺伝子の融合をもつ大腸がん,あるいは,Rspo3遺伝子を過剰に発現する非小細胞肺がんの異種移植による腫瘍の形成を抑制する.現在,転移性の大腸がんに対して第1相の臨床試験が行われている.
OMP-18R5はFz7に対するモノクローナル抗体として作製されたが,くわえて,Fz1,Fz2,Fz5,Fz8に対する認識能をもち,βカテニン経路を阻害することにより種々のヒトのがん細胞の異種移植による腫瘍の形成を抑制する42).現在,膵がん,転移性乳がん,非小細胞肺がんに対し第1相の臨床試験が行われている.
OTSA101は放射性同位元素により標識したFz10に対するモノクローナル抗体で,Fz10が滑膜肉腫において特異的に過剰に発現することから,現在,滑膜肉腫に対し第1相の臨床試験が行われている43).
βカテニンを含む複合体に対する阻害剤:PRI-724はβカテニンと転写共役因子であるCBPとの結合を阻害する低分子化合物で,βカテニンに依存的な遺伝子の発現を阻害する44).現在,βカテニンを含む複合体に対する阻害剤としては唯一,急性骨髄性白血病,慢性骨髄性白血病,大腸がん,膵がんに対し第1相および第2相の臨床試験が行われている.
βカテニン非依存性経路を標的にした抗がん剤:Cirmtuzumab(UC-961)はRor1に対するモノクローナル抗体で,Wnt5a-Ror1シグナル伝達経路を阻害することにより,Ror1遺伝子を過剰に発現する慢性リンパ性白血病細胞の増殖を抑制する45,46).現在,慢性リンパ性白血病に対し第1相の臨床試験が行われている.Foxy5はWnt5aの類似ペプチドで,Wnt5aシグナルのアゴニストとして作用する.さきに述べたように,Wnt5aはある種のがんにおいて抑制的に作用するが,実際,マウスモデルにおいてFoxy5は乳がん細胞の肺および肝転移を抑制する47).また,Wnt5a遺伝子の低発現は乳がん,大腸がん,前立腺がんにおいて予後不良と相関することから48-50),現在,これらのがんに対して第1相の臨床試験が行われている.しかし,Wnt5aはβカテニン非依存性経路を介してがんに促進的に作用することもあるので,現状では,Wnt5aのアゴニストが抗腫瘍効果を発揮するがんの明確な識別は困難である.現在,これらを識別するためのマーカー遺伝子の探索が試みられている51).
おわりに
35年以上におよぶWntの研究の結果,Wntシグナル伝達経路の異常がヒトの多様ながんに関与することが明らかにされてきた.しかし,Wntシグナルが毛包や腸管をはじめとする多くの正常な組織幹細胞の維持,増殖,分化にきわめて重要な役割をもつことから,Wntシグナルを標的にした阻害剤にはつねに深刻な副作用が懸念される.そのため,主要なWntシグナル伝達経路の構成タンパク質にくわえ,がんに特異的に機能する新たなWnt関連タンパク質やシグナル伝達経路を同定する試みは,今後,ますます重要になると考えられる.また,既存の分子標的薬や抗腫瘍薬と組み合わせてWntシグナルを阻害する試みも,副作用の回避につながるかもしれない.
最近になり,Wntシグナルはがん細胞そのものにくわえ,周囲のがん微小環境に対しても多様なはたらきをもつことが示されている.とくに,腫瘍免疫に対する作用は,今後,ますます注目されるだろう.実際,Wnt-βカテニンシグナルの活性化が腫瘍においてCD8陽性T細胞の排除にはたらくなど,抗腫瘍免疫を抑制する可能性が示唆されており52),Wntシグナル伝達経路の阻害剤とがん免疫療法との併用による治療効果の改善も期待される.
文 献
- Nusse, R. & Varmus, H. E.: Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell, 31, 99-109 (1982)[PubMed]
- Steinhart, Z. & Angers, S.: Wnt signaling in development and tissue homeostasis. Development, 145, dev146589 (2018)[PubMed]
- Kikuchi, A., Yamamoto, H. & Sato, A.: Selective activation mechanisms of Wnt signaling pathways. Trends Cell Biol., 19, 119-129 (2009)[PubMed]
- Nishita, M., Enomoto, M., Yamagata, K. et al.: Cell/tissue-tropic functions of Wnt5a signaling in normal and cancer cells. Trends Cell Biol., 20, 346-354 (2010)[PubMed]
- Langton, P. F., Kakugawa, S. & Vincent, J. P.: Making, exporting, and modulating Wnts. Trends Cell Biol., 26, 756-765 (2016)[PubMed]
- Kikuchi, A. & Yamamoto, H.: Tumor formation due to abnormalities in the β-catenin-independent pathway of Wnt signaling. Cancer Sci., 99, 202-208 (2008)[PubMed]
- Niehrs, C.: The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol., 13, 767-779 (2012)[PubMed]
- De lau, W., Peng, W. C., Gros, P. et al.: The R-spondin/Lgr5/Rnf43 module: regulator of Wnt signal strength. Genes Dev., 28, 305-316 (2014)[PubMed]
- Nusse, R. & Clevers, H.: Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell, 169, 985-999 (2017)[PubMed]
- Wiese, K. E., Nusse, R. & Van amerongen, R.: Wnt signalling: conquering complexity. Development. 145, dev165902 (2018)[PubMed]
- Polakis, P.: The many ways of Wnt in cancer. Curr. Opin. Genet. Dev., 17, 45-51 (2007)[PubMed]
- Matano, M., Date, S., Shimokawa, M. et al.: Modeling colorectal cancer using CRISPR-Cas9-mediated engineering of human intestinal organoids. Nat. Med., 21, 256-262 (2015)[PubMed]
- Satoh, S., Daigo, Y., Furukawa, Y. et al.: AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus-mediated transfer of AXIN1. Nat. Genet., 24, 245-250 (2000)[PubMed]
- Liu, W., Dong, X., Mai, M. et al.: Mutations in AXIN2 cause colorectal cancer with defective mismatch repair by activating β-catenin/TCF signalling. Nat. Genet., 26, 146-147 (2000)[PubMed]
- Wu, J., Jiao, Y., Dal Molin, M. et al.: Whole-exome sequencing of neoplastic cysts of the pancreas reveals recurrent mutations in components of ubiquitin-dependent pathways. Proc. Natl. Acad. Sci. USA, 108, 21188-21193 (2011)[PubMed]
- Giannakis, M., Hodis, E., Jasmine Mu, X. et al.: RNF43 is frequently mutated in colorectal and endometrial cancers. Nat. Genet., 46, 1264-1266 (2014)[PubMed]
- Assie, G., Letouze, E., Fassnacht, M. et al.: Integrated genomic characterization of adrenocortical carcinoma. Nat. Genet., 46, 607-612 (2014)[PubMed]
- Seshagiri, S., Stawiski, E. W., Durinck, S. et al.: Recurrent R-spondin fusions in colon cancer. Nature, 488, 660-664 (2012)[PubMed]
- Jiang, X., Hao, H. X., Growney, J. D. et al.: Inactivating mutations of RNF43 confer Wnt dependency in pancreatic ductal adenocarcinoma. Proc. Natl. Acad. Sci. USA, 110, 12649-12654 (2013)[PubMed]
- Koo, B. K., van Es, J. H., van den Born, M. et al.: Porcupine inhibitor suppresses paracrine Wnt-driven growth of Rnf43;Znrf3-mutant neoplasia. Proc. Natl. Acad. Sci. USA, 112, 7548-7550 (2015)[PubMed]
- Huang, C. L., Liu, D., Nakano, J. et al.: Wnt5a expression is associated with the tumor proliferation and the stromal vascular endothelial growth factor: an expression in non-small-cell lung cancer. J. Clin. Oncol., 23, 8765-8773 (2005)[PubMed]
- Kurayoshi, M., Oue, N., Yamamoto, H. et al.: Expression of Wnt-5a is correlated with aggressiveness of gastric cancer by stimulating cell migration and invasion. Cancer Res., 66, 10439-10448 (2006)[PubMed]
- Yamamoto, H., Oue, N., Sato, A. et al.: Wnt5a signaling is involved in the aggressiveness of prostate cancer and expression of metalloproteinase. Oncogene, 29, 2036-2046 (2010)[PubMed]
- Kremenevskaja, N., von Wasielewski, R., Rao, A. S. et al.: Wnt-5a has tumor suppressor activity in thyroid carcinoma. Oncogene, 24, 2144-2154 (2005)[PubMed]
- Bitler, B. G., Nicodemus, J. P., Li, H. et al.: Wnt5a suppresses epithelial ovarian cancer by promoting cellular senescence. Cancer Res., 71, 6184-6194 (2011)[PubMed]
- Liang, H., Chen, Q., Coles, A. H. et al.: Wnt5a inhibits B cell proliferation and functions as a tumor suppressor in hematopoietic tissue. Cancer Cell, 4, 349-360 (2003)[PubMed]
- Niehrs, C.: Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene, 25, 7469-7481 (2006)[PubMed]
- Sheng, S. L., Huang, G., Yu, B. et al.: Clinical significance and prognostic value of serum Dickkopf-1 concentrations in patients with lung cancer. Clin. Chem., 55, 1656-1664 (2009)[PubMed]
- Li, S., Qin, X., Liu, B. et al.: Dickkopf-1 is involved in invasive growth of esophageal cancer cells. J. Mol. Histol., 42, 491-498 (2011)[PubMed]
- Sato, N., Yamabuki, T., Takano, A. et al.: Wnt inhibitor Dickkopf-1 as a target for passive cancer immunotherapy. Cancer Res., 70, 5326-5336 (2010)[PubMed]
- Kimura, H., Fumoto, K., Shojima, K. et al.: CKAP4 is a Dickkopf1 receptor and is involved in tumor progression. J. Clin. Invest., 126, 2689-2705 (2016)[PubMed]
- Shinno, N., Kimura, H., Sada, R. et al.: Activation of the Dickkopf1-CKAP4 pathway is associated with poor prognosis of esophageal cancer and anti-CKAP4 antibody may be a new therapeutic drug. Oncogene, 37, 3471-3484 (2018)[PubMed]
- Kajiwara, C., Fumoto, K., Kimura, H. et al.: p63-Dependent Dickkopf3 expression promotes esophageal cancer cell proliferation via CKAP4. Cancer Res., 78, 6107-6120 (2018)[PubMed]
- Matsumoto, S., Fujii, S., Sato, A. et al.: A combination of Wnt and growth factor signaling induces Arl4c expression to form epithelial tubular structures. EMBO J., 33, 702-718 (2014)[PubMed]
- Fujii, S., Matsumoto, S., Nojima, S. et al.: Arl4c expression in colorectal and lung cancers promotes tumorigenesis and may represent a novel therapeutic target. Oncogene, 34, 4834-4844 (2015)[PubMed]
- Fujii, S., Shinjo, K., Matsumoto, S. et al.: Epigenetic upregulation of ARL4C, due to DNA hypomethylation in the 3'-untranslated region, promotes tumorigenesis of lung squamous cell carcinoma. Oncotarget, 7, 81571-81587 (2016)[PubMed]
- Hu, Q., Masuda, T., Sato, K. et al.: Identification of ARL4C as a peritoneal dissemination-associated gene and its clinical significance in gastric cancer. Ann. Surg. Oncol., 25, 745-753 (2018)[PubMed]
- Harada, T., Matsumoto, S., Hirota, S. et al.: Chemically modified antisense oligonucleotide against ARL4C inhibits primary and metastatic liver tumor growth. Mol. Cancer Ther., DOI: 10.1158/1535-7163.MCT-18-0824[PubMed]
- Jiang, X., Hao, H. X., Growney, J. D., et al.: Inactivating mutations of RNF43 confer Wnt dependency in pancreatic ductal adenocarcinoma. Proc. Natl. Acad. Sci, USA, 110, 12649-12654 (2013)[PubMed]
- van de Wetering, M., Francies, H. E., Francis, J. M. et al.: Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell, 161, 933-945 (2015)[PubMed]
- Le, P. N., Mcdermott, J. D. & Jimeno, A.: Targeting the Wnt pathway in human cancers: therapeutic targeting with a focus on OMP-54F28. Pharmacol. Ther., 146, 1-11 (2015)[PubMed]
- Gurney, A., Axelrod, F., Bond, C. J. et al.: Wnt pathway inhibition via the targeting of Frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc. Natl. Acad. Sci. USA, 109, 11717-11722 (2012)[PubMed]
- Nielsen, T. O., Poulin, N. M. & Ladanyi, M.: Synovial sarcoma: recent discoveries as a roadmap to new avenues for therapy. Cancer Discov., 5, 124-134 (2015)[PubMed]
- Gang, E. J., Hsieh, Y. T., Pham, J. et al.: Small-molecule inhibition of CBP/catenin interactions eliminates drug-resistant clones in acute lymphoblastic leukemia. Oncogene, 33, 2169-2178 (2014)[PubMed]
- Khan, A. S., Hojjat-farsangi, M., Daneshmanesh, A. H. et al.: Dishevelled proteins are significantly upregulated in chronic lymphocytic leukaemia. Tumour Biol., 37, 11947-11957 (2016)[PubMed]
- Choi, M. Y., Widhopf, G. F., Wu, C. C. et al.: Pre-clinical specificity and safety of UC-961, a first-in-class monoclonal antibody targeting ROR1. Clin. Lymphoma Myeloma Leuk., 15, S167-S1659 (2015)[PubMed]
- Safholm, A., Tuomela, J., Rosenkvist, J. et al.: The Wnt-5a-derived hexapeptide Foxy-5 inhibits breast cancer metastasis in vivo by targeting cell motility. Clin. Cancer Res., 14, 6556-6563 (2008)[PubMed]
- Dejmek, J., Dejmek, A., Safholm, A. et al.: Wnt-5a protein expression in primary dukes B colon cancers identifies a subgroup of patients with good prognosis. Cancer Res., 65, 9142-9146 (2005)[PubMed]
- Dejmek, J., Leandersson, K., Manjer, J. et al.: Expression and signaling activity of Wnt-5a/discoidin domain receptor-1 and Syk plays distinct but decisive roles in breast cancer patient survival. Clin. Cancer Res. 11, 520-528 (2005)[PubMed]
- Syed Khaja, A. S., Helczynski, L., Edsjo, A. et al.: Elevated level of Wnt5a protein in localized prostate cancer tissue is associated with better outcome. PLoS One, 6, e26539 (2011)[PubMed]
- Mehdawi, L. M., Prasad, C. P., Ehrnstrom, R. et al.: Non-canonical WNT5A signaling up-regulates the expression of the tumor suppressor 15-PGDH and indues differentiation of colon cancer cells. Mol. Oncol., 10, 1415-1429 (2016)[PubMed]
- Wang, B., Tian, T., Kalland, K. H. et al.: Targeting Wnt/β-catenin signaling for cancer immunotherapy. Trends Pharmacol. Sci., 39, 648-658 (2018)[PubMed]
著者プロフィール
略歴:2009年 広島大学大学院医歯薬学総合研究科博士課程 修了,同年 大阪大学大学院医学系研究科 特任研究員を経て,2012年より同 特任助教(現 助教).
研究テーマ:発生の過程から疾患まで多岐にわたるWntシグナルの生理的な意義.
抱負:数多くある増殖因子のなかで,Wntシグナルが具体的になぜ,どのように大事なのか,生理と病理の両面からその本質にせまりたい.
菊池 章(Akira Kikuchi)
大阪大学大学院医学系研究科 教授.
研究室URL:http://www.med.osaka-u.ac.jp/pub/molbiobc/
© 2018 松本真司・菊池 章 Licensed under CC 表示 2.1 日本
